بازدارنده رقابتی
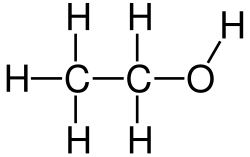
2H
5OH) بهعنوان بازدارنده رقابتی برای متانول (CH
3OH) و اتیلن گلیکول ((CH
2OH)_{
2''";}) برای آنزیم الکل دهیدروژناز در کبد عمل میکند، زمانی که به مقدار زیاد وجود داشته باشد. به همین دلیل، گاهی از اتانول بهعنوان راهی برای درمان یا پیشگیری از مسمومیت پس از بلع تصادفی این مواد شیمیایی استفاده میشود.
بازدارندگی رقابتی (انگلیسی: Competitive inhibition) نوعی مهار آنزیم است که در آن یک مادهٔ بازدارنده با مولکول طبیعیِ بستر (سوبسترا) برای نشستن در جایگاه فعال آنزیم رقابت میکند. جایگاه فعال همان بخش آنزیم است که واکنش شیمیایی در آن انجام میشود. وقتی بازدارنده وارد این جایگاه میشود، دیگر جایی برای بستر واقعی باقی نمیماند و واکنش متوقف یا کند میشود.
این فرایند را میتوان مثل یک قفل و کلید تصور کرد؛ آنزیم همان قفل است و بستر کلید درست آن. اگر یک کلید شبیه اما نادرست وجود داشته باشد (بازدارنده)، میتواند موقتاً در قفل گیر کند و جلوی کلید اصلی را بگیرد. در بازدارندگی رقابتی، اگر مقدار بستر زیاد شود، میتواند بازدارنده را کنار بزند و دوباره واکنش با سرعت طبیعی ادامه یابد.
به همین دلیل، این نوع مهار «قابل برگشت» است و در بسیاری از داروها و سموم دیده میشود. برای نمونه، بعضی داروهای کاهشدهندهٔ فشار خون یا پایینآورندهٔ کلسترول بر اساس همین اصل طراحی شدهاند تا با مهار آنزیمهای خاص، سرعت واکنشهای زیستی را کنترل کنند.[۱]
بازدارنده رقابتی به خاطر تشابه در هندسه مولکولی یا بستر برای جایگاه فعال آنزیم رقابت میکند با اشغال جایگاه فعال توسط بازدارنده از اتصال بستر با آنزیم ممانعت میکند. مهارکننده و بستر نمیتوانند همزمان با آنزیم متصل شوند.[۲] اغلب مهار کنندههای رقابتی کاملاً شبیه بستر واقعی آنزیم هستند. به عنوان مثال، داروی متوترکسات یک مهار کننده رقابتی آنزیم دی هیدروفولات ردوکتاز است که باعث کاهش دی هیدروفولات به تتراهیدروفولات میشود. شباهت بین ساختارهای دی هیدروفولات و این دارو در شکل نشان داده شده است. با غلظت زیاد بستر میتوان بر این نوع مهارکننده غلبه کرد. در برخی موارد، مهارکننده میتواند به سایتی غیر از محل اتصال سوبسترا معمولی وصل شود و یک اثر دگرریختار (آلوستریک) برای تغییر شکل محل اتصال معمولی اعمال کند.[۳]
نوع مهار آنزیم
در مهار رقابتی کاتالیز آنزیمی، اتصال یک مهارکننده مانع از اتصال مولکول هدف آنزیم، که بهعنوان بستر نیز شناخته میشود، میگردد.[۴] این عمل با مسدود کردن محل اتصال سوبسترا – یعنی جایگاه فعال – انجام میشود. Vmax حداکثر سرعت واکنش را نشان میدهد، در حالی که Km مقدار سوبسترایی است که برای رسیدن به نیمی از Vmax لازم است. Km همچنین تمایل سوبسترا برای اتصال به آنزیم را نشان میدهد.[۵] مهار رقابتی را میتوان با افزودن سوبسترای بیشتر به واکنش خنثی کرد که بخت اتصال آنزیم و بستر را افزایش میدهد. در نتیجه، مهار رقابتی تنها Km را تغییر میدهد و Vmax بدون تغییر باقی میماند. این موضوع با استفاده از نمودارهای سینتیک آنزیم، مانند مایکلایس-منتن یا لاینویور-بورک نشان داده میشود. زمانی که مهارکننده به آنزیم متصل میشود، شیب تحت تأثیر قرار میگیرد، زیرا Km از مقدار اصلی خود در واکنش افزایش یا کاهش مییابد.[۶][۷]
بیشتر مهارکنندههای رقابتی با اتصال برگشتپذیر به جایگاه فعال آنزیم عمل میکنند.[۴] به همین دلیل، بسیاری از منابع این ویژگی را بهعنوان خصوصیت تعریفکننده مهارکنندههای رقابتی بیان میکنند.[۸] با این حال، این یک سادهسازی بیش از حد گمراهکننده است، زیرا سازوکارهای مختلفی وجود دارد که در آنها آنزیم میتواند به مهارکننده یا سوبسترا متصل شود اما هرگز به هر دو بهطور همزمان متصل نمیشود.[۴] برای مثال، مهارکنندههای آلواستریک ممکن است مهار رقابتی، غیررقابتی یا غیرهمزمان را نشان دهند.[۴]
منابع
- ↑ Berg, Jeremy M. (2015). Biochemistry (8th ed.). Macmillan. ISBN 978-1464126109.
- ↑ Price NC (1979). "What is meant by 'competitive inhibition'?". Trends in Biochemical Sciences. 4 (11): N272–N273. doi:10.1016/0968-0004(79)90205-6.
- ↑ Wu P, Clausen MH, Nielsen TE (December 2015). "Allosteric small-molecule kinase inhibitors" (PDF). Pharmacology & Therapeutics. 156: 59–68. doi:10.1016/j.pharmthera.2015.10.002. PMID 26478442.
- 1 2 3 4 "Types of Inhibition". NIH Center for Translational Therapeutics. Archived from the original on 8 September 2011. Retrieved 2 April 2012.
- ↑ Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J (2000). "Functional Design of Proteins". Molecular Cell Biology (4th ed.).
- ↑ Eadie SG (1942). "The Inhibition of Cholinesterase by Physostigmine and Prostigmine". Journal of Biological Chemistry. 146: 85–93. doi:10.1016/S0021-9258(18)72452-6.
- ↑ Berg JM, Tymoczko JL, Stryer L (2002). "Appendix: Vmax and KM Can Be Determined by Double-Reciprocal Plots". Biochemistry (5th ed.).
- ↑ Ophardt C. "Virtual Chembook". Elmhurst College. Archived from the original on 17 October 2015. Retrieved 1 September 2015.