اوگلنا
| اوگلنا | |
|---|---|
 | |
| ردهبندی علمی | |
| حوزه: | |
| (طبقهبندینشده): | |
| شاخه: | |
| رده: | تاژک دار |
| راسته: | Euglenales |
| تیره: | Euglenaceae |
| سرده: | Euglena رنبرگ، ۱۸۳۰ |
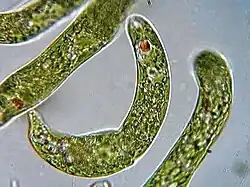
اوگلنا سردهای از جانداران تکیاختهای تاژکدار یوکاریوت است. موجود ریز تکسلولی است که همیشه زیستشناسان را متعجب میسازد، زیرا معلوم نیست که این موجود یک گیاه است یا یک جانور. چندین نوع اوگلنا وجود دارد. بعضی از آنها دارای کلروفیل بوده و مانند جلبکهای دیگر مادهسازی میکنند، اما بعضی دیگر از آنها بدون کلروفیل بوده و مانند جانوران از غذای جامد استفاده میکنند. تمامی انواع اوگلناها در داخل آب زندگی کرده و توسط حرکت تاژک درازی که دارند جابجا میشوند.[۳]
اوگلنا شناختهشدهترین عضو تاژکدارن است که بسیار مورد مطالعه قرار میگیرد. تاژکداران گروه متنوعی است که شامل حدود ۵۴ سرده و ۸۰۰ گونه است. اوگلنا دارای یک ردیاب نوری است که سلول را به سمت نور جهت می دهند و دارای لکه چشمی هستند تا فقط نور از جهت های خواص توسط ردیاب نوری جذب شود.[۴][۵] گونههای اوگلنا در آبهای تازه و دارای نمک یافت میشود. آنها اغلب در آبهای آرام درون مرز (دور از دریا) فراوان هستند، در این آبها ممکن است تعداد آنها آنقدر باشد که سطح تالابها و گودالها به رنگ سبز یا قرمز درآید.اوگلنا های دارای کلرپلاست در حضور نور فتواتوتروف(غذا ساز نوری) و در شرایط بدون نور هتروتروف(شکار گر) بوده و دیگر آغازیان را فاگوسیتوز(ذره خواری) می کنند.[۶]
در واقع این جانداران در شرایط بدون نور، با خارج کردن کلروپلاست های خود فضای کافی را برای ورود شکار خود طی فاگوسیتوز فراهم می کنند تا در این شرایط، از این راه مواد مغذی خود را تامین کنند. ضمنا در صورت بازگشت نور به شرایط محیط، از طریق ژنوم خود کلروپلاست ها را بازسازی میکند.
نکتهی حائز اهمیت درمورد جانداران واجد کلروپلاست این است که: این جانداران ارگانل مزبور را از راه "اندوسیمبیوز" و یا "کلپتوپلاستی" بدست میآورند.
در صورتی که فقط کلروپلاست به تنهایی وارد سلول جاندار شود، «کلپتوپلاستی» و اگر جاندار واجد کلروپلاست به طور کامل توسط سلول بلعیده شود، «اندوسیمبیوز» نامیده میشود. همچنین اگر این جاندار بلعیده شده پروکاریوت(مثل سیانوباکتر) باشد، اندوسیمبیوز اولیه و اگر یوکاریوت باشد، ثانویه است.
ویژگی دیگر اندوسیمبیوز برخلاف کلپتوپلاستی ، ذخیره شدن اطلاعات ساخت کلروپلاست در ژنوم است؛ بنابراین جاندارانی که با اندوسیمبیوز این ارگانل را در طی فرگشت کسب کردهاند، توانایی ساخت آن را از روی ژنوم دارند ولی این توانایی برای مثال در گوسفند دریایی که کلپتوپلاستی انجام میدهد، وجود ندارد.
کلروپلاست در اوگلنا ۳ غشایی است؛ چرا که اندوسیمبیوز ثانویه انجام میدهد.
تاریخچه
مشاهدات اولیه
اولین مشاهدهٔ مستند اوگلنا توسط آنتونی ون لیوونهوک در سال ۱۶۷۴ با میکروسکوپ اولیهاش صورت گرفت. او این موجودات متحرک را در آب بارانِ راکد توصیف کرد و بهدلیل حرکت مارپیچی منحصربهفرد، آنها را "animalcules" (جانوران کوچک) نامید. لیوونهوک متوجه شد که این ارگانیسمها بهسمت نور حرکت میکنند، اما مکانیسم این پدیده را درک نکرد. در اوایل قرن نوزدهم، کریستین گوتفرید ارنبرگ در سال ۱۸۳۰ نام علمی "Euglena" را ابداع نمود. این نام از ترکیب واژههای یونانی "eu" (خوب) و "glene" (مردمک یا هسته) گرفته شده و اشارهای به لکه چشمی قرمز رنگ و برجستهاش دارد. زیستگاه اصلی اوگلنا، آبهای شیرینِ غنی از مواد آلی نظیر برکهها، نهرهای کمجریان و باتلاقهاست، اما گونههایی مانند Euglena marina در آبهای شور ساحلی نیز یافت میشوند.[۷]
مشاهدهگران اولیه در طبقهبندی اوگلنا دچار سردرگمی بودند. گیاهشناسانی مانند ارنبرگ آن را بهدلیل داشتن کلروپلاست و توانایی فتوسنتز، در گروه جلبکهای سبز قرار دادند و اصطلاح "اوگلنوفیتا" را رواج دادند. در مقابل، جانورشناسانی چون اِدموند سِنتوینسِنت گِرَن در ۱۸۴۱، با استناد به حرکت فعال توسط تاژک و توانایی تغذیه هتروتروفیک در تاریکی، آن را یک تاژکدار جانوری ("اوگلنوزوآ") طبقهبندی کردند. این دوگانگی تا قرن بیستم ادامه یافت. شکل ظاهری اوگلنا بسیار متغیر است: طول آن از ۱۵ میکرومتر در Euglena minima تا ۵۰۰ میکرومتر در Euglena oxyuris متغیر بوده و عموماً دوکی شکل با انتهای جلویی (قدامی) پهنتر و انتهای عقبی (خلفی) نوکتیز است. حرکت مارپیچی آنها ناشی از چرخش تاژک منفرد (گاهی دو تاژک) حول محور طولی سلول است که با کمک پلیکول انعطافپذیر، امکان مانور دقیق بهسمت نور را فراهم میکند.[۸]
لکه چشمی (استیگما) یک ساختار کلیدی در مشاهدات اولیه بود. این اندامک، تودهای از رنگدانههای کاروتنوئیدی (مثل آستاگزانتین) است که در سیتوپلاسم، نزدیک پایه تاژک قرار دارد. استیگما بهعنوان یک "سایهانداز" نوری عمل میکند: با چرخش سلول، سایهٔ آن بر روی یک گیرنده نوری حساس به نور آبی در پایه تاژک (پارافلاژلار بادی) میافتد و جهت منبع نور را مشخص مینماید. این مکانیسم به اوگلنا امکان فتوتروپیسم مثبت (حرکت بهسوی نور) را میدهد. در شرایط نور مطلوب، تجمع انبوه اوگلنا در سطح آب میتواند منجر به شکوفایی جلبکی قابل رؤیت شود؛ Euglena viridis آب را سبز و Euglena sanguinea با تولید هِماتوکروم (رنگدانه قرمز)، آب را قرمز میکند. برخی گونهها مانند E. sanguinea توکسینهایی (مثل اوگلنیفریسین) تولید میکنند که برای آبزیان مضر است.[۹]
مطالعات اولیه روی تغذیه، یکی دیگر از جنبههای شگفتانگیز را نشان داد: اوگلنا در شرایط تاریکی، کلروپلاست خود را بهتدریج از دست داده و به رنگ سفید درمیآید. این شکل بیرنگ میتواند با جذب مستقیم مواد آلی محلول (ساپروتروفی) یا فاگوسیتوز ذرات ریز، زنده بماند. این پدیده از دست دادن برگزایی (اپیپلاستیدی) نامیده میشود و نشاندهندهٔ انعطاف متابولیک عمیق اوگلناست. مشاهدهٔ واکوئل انقباضی که بهطور ریتمیک منقبض میشود نیز از یافتههای اولیه بود؛ این اندامک برای دفع آب اضافی در محیطهای هیپوتونیک (مانند آب شیرین) ضروری است.[۱۰]
طبقهبندی اخیر
بر اساس دادههای مولکولی (توالییابی rRNA 18S و ژنهای پروتئینی)، اوگلنا در حوزهٔ یوکاریوتها، فرمانرو اگزکاواتا، شاخهٔ یوگلنوزوا (Euglenozoa) و ردهٔ یوگلنوئیدا (Euglenoidea) جای میگیرد. شاخهٔ یوگلنوزوا شامل سه گروه اصلی است: ۱) یوگلنوئیدهای فتوسنتزی و غیرفتوسنتزی (مانند اوگلنا) ۲) کینتوپلاستیدها (شامل تریپانوزوماهای بیماریزا) ۳) دیپلونمیدها. ویژگی مشترک آنها، ساختارهای تاژکی خاص (پارافلاژلار بادی)، ریبوزومهای بزرگ با RNA خاص، و دیانای میتوکندری با آرایش ماکسیسرکل (شبکهای) است. خویشاوندی نزدیک اوگلنا با تریپانوزوما (عامل بیماریهای خواب آفریقایی و شاگاس) نشاندهندهٔ منشأ تکاملی مشترک است، اگرچه مسیرهای تکاملی متفاوتی را پیمودهاند.[۱۱]
منشأ کلروپلاست اوگلنا نتیجهٔ یک اندوسیمبیوز ثانویه است. حدود ۲۰۰ میلیون سال پیش، یک یوگلنوئید غیرفتوسنتزی، یک جلبک سبز (از گروه کلرافیته) را بلعید اما آن را هضم نکرد. این جلبک طی تکامل به یک اندامک فتوسنتزی تبدیل شد. شواهد این فرآیند در سه لایه غشایی کلروپلاست اوگلنا دیده میشود: دو لایهٔ داخلی متعلق به غشای کلروپلاست جلبک بلعیده شده و لایهٔ سوم ناشی از غشای واکوئل غذایی میزبان است. تحلیل ژنوم نشان میدهد ژنهای کلیدی کلروپلاست به هستهٔ میزبان منتقل شدهاند (انتقال ژن افقی)، برخلاف کلپتوپلاستی در برخی آغازیان دیگر که کلروپلاست را بهطور موقت "دزدیده" و جایگزین میکنند.[۱۲]
طبقهبندی مدرن اوگلنا بیش از ۱۰۰۰ گونهٔ توصیفشده را شامل میشود که بر اساس ریختشناسی، زیستشناسی مولکولی و شیوه تغذیه به زیرگروههایی تقسیم میشوند:
اوگلنوفیتا (Euglenophyta): گونههای فتوسنتزی دارای کلروپلاست پایدار (مثل E. gracilis, E. mutabilis).
آستازیا (Astasia): گونههای بیرنگ و فاقد کلروپلاست که اجباریاً هتروتروف هستند (مثل Astasia longa که قبلاً Euglena longa نام داشت).
خانواده فاکوس (Phacidae): گونههای دارای پلیکول ضخیم و سختتر، قادر به تغییر شکل محدود. چالشهای اصلی طبقهبندی شامل تغییرپذیری ریختی (پلیمرفیسم) در پاسخ به محیط، فقدان دیوارهٔ سلولی سخت که مطالعهٔ نمونههای خشک شده را مشکل میکند، و توانایی برخی گونهها در تغییر حالت تغذیهای است که مرز بین فتواتوتروفی و هتروتروفی را محو میکند.[۱۳]
توالییابی ژنومهای کامل (مثل ژنوم Euglena gracilis در سال ۲۰۱۵) انقلابی در درک روابط فیلوژنتیک ایجاد کرده است. این دادهها تأیید میکنند که کلروپلاست اوگلنا از جلبکهای سبز (نه جلبکهای قرمز یا سیانوباکتریها) منشأ گرفته و همچنین حضور ژنهای باکتریایی متعدد در ژنوم هستهای را نشان میدهند که احتمالاً حاصل انتقال ژن افقی اضافی هستند. این ژنها ممکن است در تطابق متابولیک اوگلنا با محیطهای متنوع نقش داشته باشند.[۱۴]
فرم و عملکرد
پلیکول (Pellicle) ساختار تعیینکنندهٔ شکل اوگلناست. این پوشش، دیوارهٔ سلولی سفت نیست، بلکه از ۸۰-۲۰ نوار پروتئینی (استرپها) تشکیل شده که بهطور مارپیچی دور سلول پیچیدهاند. هر نوار از ریزلولهها و رشتههای پروتئینی (مثل آرتیکولین) ساخته شده که به آن استحکام و در عین حال انعطافپذیری میدهد. این ساختار امکان حرکت یوگلنوئیدی (Euglenoid movement یا metaboly) را فراهم میکند: سلول میتواند با لغزش استرپها روی هم، شکل خود را بهطور فعال از دوکی به کروی یا کرممانند تغییر دهد و از میان ذرات ریز یا محیطهای چسبنده عبور کند. این حرکت مستقل از تاژک است و توسط فیلامنتهای اکتین-میوزین در زیر پلیکول کنترل میشود.
حرکت تاژکی مکانیسم اصلی جابجایی است. تاژک منفرد (در بیشتر گونهها) از ساختار میکروتوبولی استاندارد ۹+۲ تشکیل شده و از درون یک حفرهٔ قدامی به نام مخزن (Reservoir) خارج میشود. حرکت آن بهصورت چرخشی و مارپیچی است که یک گرداب هیدرودینامیک در پشت سلول ایجاد کرده و پیشرانش رو به جلو ایجاد میکند. انرژی این حرکت از هیدرولیز ATP در کینتوزوم (ریزهستهٔ پایهای تاژک) تأمین میگردد. یک تاژک ثانویه کوتاهتر ممکن است وجود داشته باشد که در مخزن باقی میماند و نقشی در حرکت ندارد. جهتگیری تاژک توسط گیرندهٔ نوری پارافلاژلار (PFR) که در مجاورت استیگما قرار دارد، تنظیم میشود.[۱۵]
اوگلنا یک میکسوتروف (Mixotroph) ایدهآل است و بسته به شرایط محیطی، متابولیسم خود را تغییر میدهد:
فتواتوتروفی: در نور، فتوسنتز با استفاده از کلروفیل a و b (مشابه گیاهان) و کاروتنوئیدهای فرعی (مثل دیادینوکسانتین) انجام میدهد. محصول فتوسنتز بهصورت پارامیلون، یک پلیساکارید ذخیرهای β-1,3 گلوکان، در دانههای بزرگ در سیتوپلاسم ذخیره میشود. پارامیلون توسط آنزیم پارامیلاز تجزیه میگردد.
هتروتروفی: در تاریکی یا کمنوری، مواد آلی محلول (مانند استات، اتانول، اسیدهای آمینه) را مستقیماً از طریق پروتئینهای ناقل غشایی جذب میکند. همچنین برخی گونهها قادر به فاگوسیتوز باکتریها یا ذرات ریز با تشکیل واکوئلهای غذایی هستند.
اُسموتروفی: جذب مواد مغذی محلول در محیط بهویژه در گونههای بیرنگ.[۱۶]
واکنشهای محیطی:
فتوتروپیسم مثبت: جذب بهسوی نور با شدت متوسط (۴۰۰–۵۰۰ نانومتر، نور آبی-سبز) با دقت بالا.
فوبیفتوتروپیسم: فرار از نور شدید مستقیم (UV یا شدید سفید) برای جلوگیری از فوتواکسیداسیون و تخریب رنگدانهها.
گراوتروپیسم: پاسخ به گرانش، بهویژه در گونههای خاکزی.
کموتروپیسم: حرکت بهسمت یا دور از غلظتهای خاص مواد شیمیایی (مثل اکسیژن، مواد مغذی). این پاسخها توسط یک شبکهٔ سیگنالینگ پیچیده هماهنگ میشوند که کلسیم یونهای ثانویه، cAMP و فسفریلاسیون پروتئینها در آن نقش دارند.[۱۷]
اجزا
تاژک (Flagellum): اندامک حرکتی اولیه. دارای آرایش میکروتوبولی ۹+۲ (۹ دوبلت محیطی + ۲ سینگلت مرکزی)، پوشیده از موتیل (Mastigonemes) یا تارهای جانبی. به کینتوزوم (جایگاه تولید میکروتوبول) در پایه متصل است. حرکت آن توسط داینئینهای وابسته به ATP انجام میشود.
مخزن (Reservoir): حفرهٔ قیفیشکل در انتهای قدامی سلول که تاژک از آن خارج میشود. محل اتصال واکوئل انقباضی است.
لکه چشمی (Stigma یا Eyespot): مجموعهای از حبابهای لیپیدی حاوی رنگدانههای کاروتنوئیدی (عمدتاً β-کاروتن و دیادینوکسانتین). بهعنوان فیلتر نوری و شاتر جهتیاب عمل میکند. فاقد هرگونه عدسی یا ساختار تصویرساز است.
واکوئل انقباضی (Contractile Vacuole): سیستم دفع آب اضافی. در آب شیرین فعالتر است؛ آب را از سیتوپلاسم جمع کرده و به درون مخزن تخلیه میکند. چرخهٔ انقباض آن ۱۵-۲۰ ثانیه طول میکشید.
کلروپلاست (Chloroplast): اندامک فتوسنتزی با غشای سهلایه. حاوی تیلاکوئیدهای گروهبندیشده بهصورت ۳-۵ تایی (برخلاف گرانا در گیاهان). رنگدانهها: کلروفیل a و b، بتاکاروتن، دیادینوکسانتین، نوگیریکسانتین.
پارامیلون (Paramylon): دانههای ذخیرهای سفید-براق β-1,3 گلوکان. غیرنشاستهای، غیرفلورسنت و مقاوم به آمیلاز. در سیتوپلاسم پراکنده یا نزدیک کلروپلاست.
هسته (Nucleus): هستهٔ یوکاریوتی بزرگ با نوکلئولوس مشخص. دارای کروموزومهای دائماً کندِنسه (کوندری شده) در اینترفاز.
پلیکول (Pellicle): پوشش خارجی انعطافپذیر متشکل از نوارهای پروتئینی متحرک. حاوی پروتئینهایی مانند آرتیکولین و ایپسیلین.
میتوکندری (Mitochondrion): دارای کریستاهای لولهای. ژنوم میتوکندری (کینتوپلاست در خویشاوندانش نیست) بهصورت ماکسیسرکلهای درهمتنیده.
اجسام مویی (Muciferous Bodies): وزیکولهای زیر پلیکول حاوی موکوپلیساکارید. احتمالاً در ترشح غشاء یا تشکیل کیست نقش دارند.
تولید مثل
تولید مثل غیرجنسی اوگلنا عمدتاً از طریق تقسیم دوتایی طولی (Longitudinal Binary Fission) صورت میگیرد. این فرآیند میتوزی در شرایط مطلوب (نور، مواد مغذی کافی، دمای ۲۰-۲۵°C) هر ۶-۲۴ ساعت رخ میدهد. مراحل آن شامل: ۱. پیشتقسیم: همانندسازی DNA هستهای، کینتوزومها و کلروپلاستها. تاژک ممکن است جذب شده یا باقی بماند. ۲. تقسیم هسته: میتوز با حفظ پوشش هستهای (میتوز بسته). ۳. تقسیم سیتوپلاسم: از قطب قدامی (نزدیک مخزن) به سمت قطب خلفی پیش میرود. سلول کشیده شده و پلیکول در امتداد محور طولی تقسیم میشود. ۴. جدا شدن: دو سلول دختری تشکیل میشوند. هر سلول یک تاژک جدید میسازد (اگر تاژک مادری جذب شده بود) یا تاژک مادری به یکی منتقل شده و دیگری تاژک جدید میسازد. سلولهای دختری کوچکتر از والد هستند و طی چند ساعت به اندازهٔ طبیعی میرسند.
در شرایط استرسزا (خشکی، کمبود مواد مغذی، دمای نامناسب، آلودگی)، اوگلنا بهشکل یک کیست (Cyst) یا هاگ دیوارهدار درمیآید. فرآیند کیستزایی شامل:
- از دست دادن تاژک و گرد شدن سلول.
- ترشح یک دیوارهٔ ضخیم چندلایه (غنی از موکوپلیساکارید و اسپوروپولنین) از اجسام مویی.
- کاهش شدید متابولیسم و توقف تقسیم.
متراکمشدن سیتوپلاسم و تجمع ذخایر پارامیلون. کیستها میتوانند ماهها تا سالها در رسوبات خشک یا یخزده زنده بمانند. با بازگشت شرایط مطلوب (رطوبت، نور، مواد مغذی)، دیوارهٔ کیست حل شده، سلول بهشکل تروفوزوئیت فعال خارج میشود و تاژک جدیدی میسازد.[۱۸]
اگرچه گزارشهای تاریخی از تولید مثل جنسی (همیوغی) وجود دارد، اما هیچ شواهد مولکولی یا سیتولوژیک قانعکنندهای برای وقوع میوز، گامتزایی یا ترکیب هستهای در اوگلنا یافت نشده است. تنوع ژنتیکی در جمعیتهای اوگلنا عمدتاً از طریق جهشهای خودبهخودی و بازترکیبی در طی تقسیم میتوز (بهویژه در طولانیشدن تلومرها) ایجاد میشود. برخی مطالعات انتقال ژن افقی از باکتریها را نیز گزارش کردهاند.[۱۹]
موارد استفاده

تولید سوخت زیستی (Biofuel)
Euglena gracilis بهدلیل رشد سریع، توانایی رشد در فاضلابهای شهری/صنعتی و ذخیرهٔ بالای لیپیدهای خنثی (تا ۵۹٪ وزن خشک در شرایط استرس نیتروژن) بهعنوان منبع امیدوارکننده برای تولید بیودیزل استفاده میشود.
شرکت ژاپنی Euglena Co. در سال ۲۰۱۸ اولین کارخانهٔ تجاری تولید بیودیزل از اوگلنا را راهاندازی کرد.
پارامیلون نیز میتواند به اتانول زیستی تبدیل شود.[۲۰]
تغذیه انسانی و دامی
زیستتودهٔ اوگلنا غنی از پروتئین کامل (۵۰-۶۰٪ وزن خشک، حاوی تمام اسیدهای آمینهٔ ضروری)، ویتامینها (E, B1, B12 بالا)، اسیدهای چرب امگا-۳ (DHA، EPA) و مواد معدنی است.
پودر "یوجلنا" (Euglena) توسط شرکت ژاپنی بهعنوان سوپرفود بهصورت کپسول، پودر نوشیدنی و افزودنی غذایی عرضه میشود. ادعا میشود سیستم ایمنی و متابولیسم را تقویت میکند.
بهعنوان مکمل غذایی در آبزیپروری و خوراک دام استفاده میشود.[۲۱]
تصفیه فاضلاب (Wastewater Treatment)
اوگلنا قادر به جذب و متابولیسم مقادیر بالای نیتراتها، فسفاتها، آمونیاک و فلزات سنگین (مثل Cd, Pb) از فاضلابهای کشاورزی، صنعتی و شهری است (فیتورمدیشن).
همزمان با تصفیه، زیستتودهٔ باارزش تولید میکند که برای سوخت زیستی یا خوراک قابل برداشت است.
سیستمهای فوتوبیوراکتور اختصاصی برای کشت اوگلنا در فاضلاب توسعه یافتهاند.[۲۲]
جذب دیاکسید کربن (CO2 Sequestration):
ظرفیت فتوسنتزی بالا (رشد سریع) اوگلنا آن را برای جذب کربن در دودکشهای صنعتی یا سیستمهای بسته مناسب میکند.
پروژههایی مانند "Euglena Bio-factory" هدف جذب CO2 و تولید محصولات باارزش را دنبال میکنند.[۲۳]
زیستفناوری و پژوهش
زیستحسگرها (Biosensors): استفاده از تغییرات حرکت، فلورسانس کلروفیل یا بیان ژن اوگلنا برای تشخیص آلایندههای آب (سموم، فلزات سنگین، علفکشها).
مدل تحقیقاتی: مطالعهٔ مکانیسمهای پایهٔ زیستشناسی سلولی (حرکت تاژکی، فتوتاکسی، اندوسیمبیوز، انعطاف متابولیک).
فضا: ناسا و آژانس فضایی اروپا (ESA) از اوگلنا در سیستمهای پشتیبانی حیات زیستی (BLSS) و تحقیقات گراوتروپیسم در میکروگرانش استفاده کردهاند (مثلاً در پروژههای "حیات در فضای بسته").
تولید ترکیبات باارزش: تولید ویتامینها، اسیدهای آمینه، آنتیاکسیدانها (مثل اسید اسکوربیک، توکوفرول) و حتی آنتیبیوتیکها (مثل اوگلنینول) در کشتهای تغذیهشده.[۲۴]
کاربردهای عمدهٔ اوگلنا در صنعت و پژوهش
کاربرد گونه نمونه محصول/فایده وضعیت توسعه سوخت زیستی Euglena gracilis بیودیزل، بیواتانول از لیپیدها و پارامیلون تجاری (ژاپن)
تغذیه انسانی Euglena gracilis پودر پروتئینی، مکمل ویتامینی، اسیدهای چرب تجاری (ژاپن، اروپا)
تصفیه فاضلاب E. mutabilis حذف نیترات/فسفات، تولید زیستتوده همزمان پایلوت و نیمهصنعتی
جذب CO2 صنعتی E. gracilis کاهش انتشار گازهای گلخانهای تحقیقاتی-پایلوت
زیستحسگرها E. gracilis تشخیص آلودگی آب بر اساس تغییرات رفتاری/زیستی تحقیقاتی [۲۵]
جستارهای وابسته
منابع
- مشارکتکنندگان ویکیپدیا. «Euglena». در دانشنامهٔ ویکیپدیای انگلیسی، بازبینیشده در ۱۹ اکتبر ۲۰۱۷.
- ↑ Adl, SM; Simpson, AG; Lane, CE; Lukeš, J; Bass, D (2012). "The Revised Classification of Eukaryotes". Journal of Eukaryotic Microbiology. 59 (5): 429–493. doi:10.1111/j.1550-7408.2012.00644.x. PMC 3483872. PMID 23020233.
- ↑ Guiry, MD; Guiry, GM. "Algaebase Taxonomy Browser". Algaebase. National University of Ireland. Retrieved 2015-05-11.
- ↑ گیاهان بیگل / مترجم احد اشگریز. تبریز. سازمان تحقیقات، آموزش و ترویج کشاورزی. ناشر: حکمت. ؟۱۳۶ .
- ↑ "The Euglenoid Project: Alphabetic Listing of Taxa". The Euglenoid Project. Partnership for Enhancing Expertise in Taxonomy. Archived from the original on 23 February 2017. Retrieved Sep 20, 2014.
- ↑ "The Euglenoid Project for Teachers". The Euglenoid Project for Teachers. Partnerships for Enhancing Expertise in Taxonomy. Archived from the original on 23 February 2017. Retrieved Sep 20, 2014.
- ↑ Wolosski, Konrad. "Phylum Euglenophyta". In John, David M.; Whitton, Brian A.; Brook, Alan J. (eds.). The Freshwater Algal Flora of the British Isles: an Identification Guide to Freshwater and Terrestrial Algae. p. 144. ISBN 978-0-521-77051-4.
- ↑ Leedale, G. F. (1978). Phylogenetic Criteria in Euglenoid Flagellates. BioSystems, 10(1-2), 183–194. doi:10.1016/0303-2647(78)90040-0
- ↑ Schnepf, E. (1993). From Extrusomes to Trichocysts: A Phylogenetic View. European Journal of Protistology, 29(4), 359–370. doi:10.1016/S0932-4739(11)80400-2
- ↑ Barsanti, L., & Gualtieri, P. (2009). Euglena: The Experimental Organism. In Encyclopedia of Life Sciences (ELS). John Wiley & Sons, Ltd. doi:10.1002/9780470015902.a0001966.pub2
- ↑ Leander, B. S., & Farmer, M. A. (2001). Comparative Morphology of the Euglenid Pellicle. II. Diversity of Strip Substructure. Journal of Eukaryotic Microbiology, 48(2), 202–217. doi:10.1111/j.1550-7408.2001.tb00304.x
- ↑ Adl, S. M., et al. (2019). Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. Journal of Eukaryotic Microbiology, 66(1), 4–119. doi:10.1111/jeu.12691
- ↑ Keeling, P. J. (2010). The Endosymbiotic Origin, Diversification and Fate of Plastids. Philosophical Transactions of the Royal Society B: Biological Sciences, 365(1541), 729–748. doi:10.1098/rstb.2009.0103
- ↑ Zakryś, B., et al. (2017). An Integrative Approach to the Genus Euglena (Euglenaceae): Species Discrimination and Phylogenetic Relationships. Phycologia, 56(3), 342–358. doi:10.2216/16-48.1
- ↑ Ebenezer, T. E., et al. (2019). Transcriptome, Proteome and Draft Genome of Euglena gracilis. BMC Biology, 17(1), 11. doi:10.1186/s12915-019-0626-8
- ↑ Suzaki, T., & Williamson, R. E. (1980). Euglenoid Movement in Euglena fusca: Evidence for Sliding Between Pellicular Strips. Protoplasma, 124(3), 137–146. doi:10.1007/BF01290723
- ↑ Buetow, D. E. (1968). The Biology of Euglena (Vol. 1). Academic Press. ISBN 978-0121388016
- ↑ Dieckmann, C. (1995). Gravitaxis in Euglena. Advances in Space Research, 17(6-7), 73–82. doi:10.1016/0273-1177(95)00622-N
- ↑ Triemer, R. E., & Zakryś, B. (1998). Ultrastructure of Cyst Formation in Euglena. Journal of Eukaryotic Microbiology, 45(1), 43–50. doi:10.1111/j.1550-7408.1998.tb05067.x
- ↑ Haider, S., et al. (2002). Search for Evidence of Sexual Reproduction in Euglena gracilis. European Journal of Protistology, 38(3), 313–318. doi:10.1078/0932-4739-00855
- ↑ Grima, E. M., et al. (2013). Microalgae for Biofuels Production: Current Status and Future Prospects. Renewable and Sustainable Energy Reviews, 26, 241–264. doi:10.1016/j.rser.2013.05.019
- ↑ Shimada, R., et al. (2009). Effects of Euglena gracilis Intake on Human Health. Journal of Nutritional Science and Vitaminology, 55(Supplement), S100. (Conference Abstract).
- ↑ Ahmed, S. F., et al. (2014). Phycoremediation of Wastewater Using Euglena. International Journal of Environmental Science and Technology, 11(6), 1807–1816. doi:10.1007/s13762-013-0418-6
- ↑ Kumar, K., et al. (2010). Microalgae as a Renewable Source of Energy: A Niche Opportunity. Journal of Chemical Technology & Biotechnology, 85(1), 11–18. doi:10.1002/jctb.2287
- ↑ Gissibl, A., et al. (2021). Euglena gracilis as a Model Organism in Biomedical Research. Microorganisms, 9(9), 1888. doi:10.3390/microorganisms9091888
- ↑ Euglena Co., Ltd. (2023). Sustainability Report 2023. Retrieved from https://www.euglena.jp/en/sustainability/report/ (Official Company Report)
پیوند به بیرون
- The Euglenoid Project
- Tree of Life web project: Euglenida
- Protist Images: Euglena
- Euglena at Droplet - Microscopy of the Protozoa
- Images and taxonomy
- Constantopoulos, George; Bloch, Konrad (1967). "Effect of Light Intensity on the Lipid Composition of Euglena gracilis". The Journal of Biological Chemistry. 242 (15): 3538–42.