کلاوستروم
| عایق | |
|---|---|
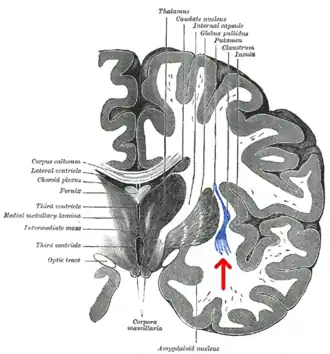 برش کرونال از مخ انسان. عایق با پیکان مشخص شده است. | |
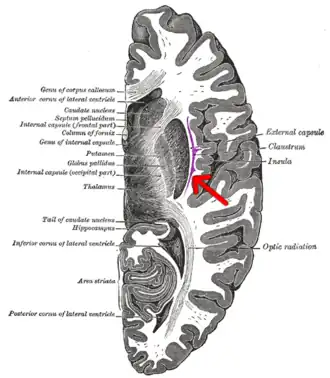 برش عرضی از مخ انسان. (سمت قدامی در بالا قرار دارد) عایق با پیکان مشخص شده است. | |
| جزئیات | |
| بخشی از | مغز |
| سرخرگها | سرخرگ مغزی میانی |
| شناسهها | |
| MeSH | D000079482 |
| نورونیمز | 252 |
| شناسه نورولکس | birnlex_1522 |
| TA98 | A14.1.09.421 |
| TA2 | 5535 |
| FMA | 67440 |
عایِقِ مغز یا کلاوستروم[۱] (انگلیسی: Claustrum) بخشی نازک و باریک از مغز است که بین قشر مغز و هستههای عمیقتر قرار دارد. این ناحیه در هر دو نیمکرهٔ مغزی وجود دارد و با بخشهای زیادی از مغز ارتباط دارد.
به زبان ساده، عایق مغز مانند یک مرکز هماهنگکننده است که سیگنالهای ورودی را از نقاط مختلف مغز دریافت میکند و ممکن است در ترکیب و یکپارچهسازی آگاهی، توجه، و درک حسی نقش داشته باشد.
برخی پژوهشگران آن را «هادی آگاهی» نامیدهاند، چون تصور میشود به مغز کمک میکند تا اطلاعات پراکنده را در یک تجربهٔ منسجم ترکیب کند.
در تعریف علمیتر، عایق مغز لایهای نازک از نورونها و سلولهای گلیال پشتیبان در مغز است که با قشر مغز و نواحی زیرقشری از جمله آمیگدال، هیپوکامپ و تالاموس ارتباط دارد.[۲][۳] این ساختار میان قشر جزیرهای از طرف خارجی و پوتامن از طرف داخلی قرار گرفته است و بهترتیب درون کپسول خارجی و کپسول بسیار خارجی جای دارد.[۴][۲][۵] سرخرگ مغزی میانی خونرسانی به عایق مغز را تأمین میکند.[۲] این ساختار بهعنوان متراکمترین شبکهٔ متصل در مغز شناخته میشود، از همین رو فرض بر این است که در یکپارچهسازی ورودیهای گوناگون قشری مانند بینایی، شنوایی و بساوایی در قالب گشتالت نقش ایفا میکند.[۵][۶] فرضیههای دیگر بیان میکنند که عایق مغز در پردازش اهمیتبرانگیز دخالت دارد تا توجه را به محرکهای رفتاری مهم در میان نوفههای (نویزهای) زمینهای معطوف کند.[۷] بررسی این ساختار دشوار است، زیرا افراد دارای ضایعه در ناحیهٔ عایق مغز اندکاند و وضوح تصویربرداری عصبی پایین است.
عایق مغز از انواع گوناگونی از سلولها تشکیل شده که از نظر اندازه، شکل و ترکیب شیمیایی-عصبی با هم تفاوت دارند. تاکنون پنج نوع سلول شناسایی شدهاند که بیشترشان به نورون هرمی در قشر مغز شباهت دارند.[۸][۹] درون عایق مغز برخلاف قشر مغز، سازمان لایهای یاختهها وجود ندارد و جسم سلولی میتواند بهصورت هرمی، دوکیشکل یا دایرهای باشد.[۲] سلول اصلی موجود در این ناحیه، نورون نوع I گلژی است؛ سلولی بزرگ با دندریتهایی که دارای خارهای متعدد هستند.[۱۰][۹]
از طریق ارتباطات میاننیمکرهای، باور بر این است که عایق مغز در همزمانسازی فعالیت در بخشهای مختلف اما کارکردیِ مرتبط مغز، مانند ناحیهٔ میدان بینایی پیشانی و قشر بینایی نقش دارد.[۲][۱۱][۱۲] از این رو، گمان میرود که عایق مغز در یکپارچهسازی اطلاعات حسی گوناگون نقش دارد و شاید خودآگاهی را نیز پشتیبانی کند.[۱۱][۱۳] عملکرد پیشنهادی دیگر برای این ساختار آن است که اطلاعات مهم را از اطلاعات بیاهمیت تمایز دهد تا دومی نادیده گرفته شود.[۶][۱۳][۱۴]
اجزای قشری مرتبط با خودآگاهی شامل قشر پیشانی-جداری، قشر کمربندی و پیشگُوِه هستند. بهسبب ارتباطات فراگیر عایق مغز با این نواحی، گفته میشود که این ساختار ممکن است در توجه و خودآگاهی نقش داشته باشد. شبکههای عصبی مسئول توجه پایدار و خودآگاهی، ورودیهایی به عایق مغز ارسال میکنند و در گزارشی از یک مورد انسانی، تحریک الکتریکی نزدیک به این ساختار، وضعیت آگاهی فرد را بهصورت بازگشتپذیر مختل کرد.[۱۵]
ساختار
عایق مغز ساختاری دوجانبه از ماده خاکستری است که اندازهٔ آن حدود ۰٫۲۵٪ از قشر مغز را تشکیل میدهد و در عمق قشر جزیرهای و کپسول بسیار خارجی، و در سطح کپسول خارجی و گرههای پایهای قرار دارد.[۲]
نام لاتین این عایق، یعنی کلاوستروم، بهمعنای «پنهانشده» است و نخستینبار در سال ۱۶۷۲ شناسایی شد. توصیفهای دقیقتر از این ساختار در سدهٔ نوزدهم میلادی ارائه شد.[۲] اگرچه مرزهای کالبدشناختی منطقهایِ عایق مغز مشخص شدهاند، در تعریف دقیق حدود آن در ادبیات علمی همچنان اجماع وجود ندارد،[۱۴][۱۶][۱۷] با اینحال، در سال ۲۰۱۹ گروهی از متخصصان چارچوبی برای شناسایی ساختارهای مرتبط با عایق مغز در گونههای مختلف ارائه کردند.[۴]
ارتباطات
در نخستین بررسیهای قرن بیستم، تأکید بر ورودیها و خروجیهای قشری بود.[۱۸] با این حال، پژوهشهای بعدی نشان دادهاند که عایق مغز با نواحی قشری و زیرقشری، ارتباطات فراوانی دارد.[۱۹] مطالعات الکتروفیزیولوژی نشان دادهاند که این ساختار با هستههای تالاموسی و عقدههای قاعدهای پیوندهای گستردهای دارد، درحالیکه گزارشهای ایزو توپولوژیک نیز ارتباط عایق مغز با نواحی پیشپیشانی، پیشانی، جداری، گیجگاهی و پسسری را نشان دادهاند.[۲۰][۲۱] مطالعات بیشتری نیز به بررسی ارتباط این ساختار با مسیرهای مادهٔ سفید زیرقشری پرداختهاند. ساختارهایی همچون تاج شعاعی، دستهٔ پیشانی-پسسری و دسته قلابی از نواحی پیشانی، مرکزی، جداری و پسسری به عایق مغز فرافکنی دارند.[۲۲] ارتباطات دوسویه نیز با نواحی حرکتی، سامانه حسی-پیکری، قشر شنوایی و نواحی بینایی وجود دارد.[۱۴] در مجموع، یافتهها نشان میدهند که عایق مغز متراکمترین ساختار متصلشده بهازای حجم منطقهای در مغز است و ممکن است بهعنوان یک «گرهٔ هماهنگکننده» برای تنظیم فعالیت مدارهای مغزی عمل کند.[۲۳][۲۴] با وجود این گستردگی، بیشتر ارتباطات فرافکن و دریافتی این ساختار در همان سمت مغز (همسوی نیمکره) هستند، اگرچه تعداد محدودی ارتباط دگرسو نیز شناسایی شدهاند. شواهد اندکی دربارهٔ ارتباط این ساختار با ساقه مغز و نخاع وجود دارد.[۱۴][۱۸][۲۵] در مجموع، ارتباطات قشری و زیرقشری این ساختار نشان میدهند که عایق مغز در پردازش اطلاعات حسی و همچنین وضعیت جسمی و هیجانی جاندار نقش دارد.
ریزکالبدشناسی
ورودیهای عایق مغز بر پایهٔ نوع اطلاعات دریافتی (مدالیته) سازمان یافتهاند؛ این ورودیها شامل نواحی پردازش پیشپیشانی، بینایی، شنوایی و دستگاه عصبی پیکری هستند. همانگونه که ساختار نورونها در لایههای رِکسِد طناب نخاعی نشاندهندهٔ کارکرد آنهاست، نواحی بینایی، شنوایی و حرکتی-پیکری در عایق مغز نیز دارای نورونهایی با ویژگیهای عملکردی ویژه و مشابهاند. برای نمونه، بخشی از عایق مغز که اطلاعات بینایی را پردازش میکند («در درجهٔ نخست، رشتههای آوران مربوط به میدان بینایی پیرامونی ما را ترکیب میکند») عمدتاً از سلولهای دوچشمی تشکیل شده که «دارای میدانهای گیرندهٔ کشیده و فاقد گزینشگری جهت» هستند.[۲۶][۲۷]
تمرکز بر سامانهٔ حسی پیرامونی پدیدهای جداگانه نیست، چراکه بیشتر رشتههای آوران حسی واردشونده به عایق مغز، اطلاعات پیرامونی را انتقال میدهند. افزون بر این، عایق مغز برای هر مدالیتهٔ حسی دارای سازماندهی مکانی متمایزی است و همچنین با قشرهای پیشانی ارتباطی متراکم دارد.[۲۸][۲۹] برای نمونه، در ناحیهٔ پردازش بیناییِ عایق مغز، سازمان بینایینگر (رتینوتوپیک) وجود دارد که با نواحی ارتباطی بینایی و ناحیهٔ V1 همراستا است، مشابه با حفظ این سازمان در هستهٔ جانبی زانویی، اما با پیچیدگی کمتر.[۱۴]
درون عایق مغز، ارتباطات موضعی عمدتاً از طریق مهار دوسیناپسی پیشسو صورت میگیرد، بهطوریکه در آن، نورونهای میانجیِ دارای پاروالبومین، فعالیت نورونهای فرافکن مجاور را سرکوب میکنند.[۳۰] خودِ نورونهای میانجی از طریق سیناپس و اتصال شکاف بههم متصلاند، که این امر باعث مهار گسترده و همزمان مدارهای موضعی عایق مغز میشود. در مطالعات اخیر بر روی عایق مغز در موشها[۲۸] و خفاشها،[۳۱] مشاهده شده که نورونهای برانگیزندهٔ فرافکن به قشر مغز، سیناپسهایی در امتداد محور پیشعقب تشکیل میدهند و بهسمت نورونهایی متمایلاند که مقصدهای فرافکنی مشابهی ندارند؛ عملکرد احتمالی این وضعیت، پیوند دادن فعالیت ماژولهای آوران مختلف است.[۲۹] درمجموع، این دو نوع مدار حاکی از آناند که عایق مغز توانایی تبدیل موضعی اطلاعات ورودی متنوع از سراسر مغز را دارد.
انواع یاختهها
عایق مغز از انواع مختلفی از سلولها تشکیل شده که در اندازه، شکل و ترکیب شیمیایی-عصبی با یکدیگر تفاوت دارند.[۵] سلولهای برانگیزندهٔ موجود در این ساختار به دو ردهٔ اصلی تقسیم میشوند که بهطور متمایز به نواحی قشر مغز و زیرقشری فرافکنی دارند. نورونهای مهاری تنها ۱۰ تا ۱۵ درصد از کل نورونهای عایق مغز را تشکیل میدهند و شامل سه نوعاند که بهترتیب دارای پاروالبومین، گیرنده سوماتواستاتین نوع ۲ یا گیرنده پپتید وازواکتیو رودهای هستند و با نورونهای مهاری موجود در قشر مغز مشابهاند.[۳۲] چندین مطالعه نشان دادهاند که بهترین ویژگی ساختاریِ متمایزکنندهٔ عایق مغز، شبکهٔ آشکار رشتههای دارای پاروالبومین است که توسط سلولهای مهاری دارای این ماده ساخته میشود.[۶] در پژوهشهای نوین، از پروتئین اصلی میلین (MBP) و توکسین وبای بازگشتی نیز بهعنوان روشهای مؤثر برای شناسایی عایق مغز استفاده شده است.[۲۹][۳۳]
در موشها، رویکردهای گوناگونی برای بررسی انواع سلولهای عایق مغز بهکار رفته که شامل روشهای الکتروفیزیولوژیکی، ریختشناختی، ژنتیکی و ارتباطی (کانکتومیک) است.[۲۸][۲۹][۳۴][۳۵][۳۶] اگرچه هنوز دربارهٔ شمار دقیق انواع سلولهای برانگیزنده توافق روشنی وجود ندارد، پژوهشهای اخیر نشان دادهاند که نورونهای فرافکن به قشر و زیرقشر، احتمالاً متمایز از یکدیگرند و در ویژگیهایی چون ویژگیهای الکتروفیزیولوژیکی ذاتی، ورودیهای آوران و نیمرخهای تعدیلکننده عصبی تفاوت دارند.[۲۸][۳۴][۳۷]
کارکرد
نشان داده شده است که عایق مغز در بخشهای گستردهای از قشر مغز فعالیت دارد و این نواحی همگی با اجزای مرتبط با خودآگاهی و توجه پایدار در ارتباطاند. این امر بهسبب پیوندهای فراگیر عایق مغز با نواحی پیشانی-جداری، قشر کمربندی و تالاموسها است. توجه پایدار بهویژه از طریق ارتباطات با قشر کمربندی، قشر گیجگاهی و تالاموس امکانپذیر میشود.
فرانسیس کریک و کریستوف کوخ پیشنهاد کردهاند که عایق مغز نقشی همانند رهبر ارکستر دارد که سعی میکند کارکرد تمام ارتباطات را هماهنگ کند.[۲] این تمثیل «رهبر ارکستر» با در نظر گرفتن پیوندهای میان عایق مغز، نواحی حسی و نواحی پیشپیشانی تقویت میشود. ثابت شده که عایق مغز با قشر پیشپیشانی، نواحی بینایی، شنوایی، حسی و حرکتی پیوند دوسویه دارد. این ارتباطات بین مدالیتههای مختلف دیدی کلی از کارکرد عایق مغز ارائه میدهند. در اینجا فرض میشود که عایق مغز در «درگاهگذاریِ توجه گزینشی» نقش دارد. از طریق این فرایند درگاهگذاری، عایق مغز میتواند ورودی از این مدالیتهها را بهطور گزینشی تنظیم کرده و فرایند «تمرکز» را تسهیل کند. همچنین پیشنهاد شده که عملکردی معکوس نیز دارد؛ یعنی از طریق نرمالسازی تقسیمکننده،[۳۸] عایق مغز ممکن است در برابر برخی ورودیها مقاومت ایجاد کرده و مانع «حواسپرتی» شود.
کارکرد احتمالی
برای اینکه عایق مغز بتواند در خودآگاهی نقش داشته باشد، باید توانایی یکپارچهسازی مدالیتههای حسی و حرکتی گوناگون از نواحی مختلف قشر مغز را داشته باشد. اتصالات کالبدشناختی عایق مغز از طریق تصویربرداری پخش وزنی (DTI) بررسی شدهاند. افامآرآی (fMRI) با سنجش سطح اکسیژن خون در مغز، فعالیت نواحی خاص قشری را نشان میدهد. این اسکنها در وضعیت بیهوشی در مقایسه با بیداری در موشها، فعالیت کمتری را در عایق مغز نشان میدهند، بهویژه در پیوندهای عایق مغز با قشر پیشپیشانی (mPFC) و هسته پشتی میانی تالاموس (MD thalamus). عایق مغز با عایق مغزِ نیمکرهٔ مقابل نیز ارتباطی قوی و کارکردی دارد. همچنین با هستهٔ MD, mPFC و نواحی قشری نزدیک و دور نیز پیوند دارد.[۱۱]
تحریک الکتریکی ناحیهٔ پشتی عایق مغز در گربهها، پاسخهای برانگیزندهای در قشر بینایی ایجاد میکند. عایق مغز از نظر کالبدی در محل تلاقی تعداد زیادی از دستههای مادهٔ سفید مغز قرار دارد که برای اتصال نواحی گوناگون قشر مغز بهکار میروند. این نیز نقش مرکزیِ ادغام اطلاعات حسی و حرکتی را برای این ساختار تقویت میکند. وجود اتصال شکاف میان نورونهای بیخار (فاقد دندریتهای شاخهدار) در عایق مغز نیز نشان میدهد که این ساختار میتواند مدالیتههای مختلف را بهصورت همزمان هماهنگ کند.[۲]
مطالعات دیگری نیز نقش عایق مغز را در حافظه فضایی[۳۹] و خواب امواج آهسته مطرح کردهاند.[۴۰][۴۱]
توجه
عایق مغز توانایی متمایز کردن اطلاعات مرتبط با وظیفه از اطلاعات نامرتبط را دارد تا بتواند توجه هدایتشده ایجاد کند. این ساختار، بیشترین تراکم ارتباطی در میان رشتههای مادهٔ سفید در قشر مغز را داراست، که از نقش آن در شبکهسازی و هماهنگی میان نواحی گوناگون مغز پشتیبانی میکند.[۱۳] عایق مغز ویژگی منطقهای خاص دارد؛ یعنی اطلاعات ورودی از مراکز بینایی به نواحی خاصی از ماده خاکستری در این ساختار و قشر شنوایی فرافکنی مییابد.[۲] تحریکهای غیرمنتظره نیز عایق مغز را فعال کرده و باعث تمرکز فوری یا تخصیص کارکرد میشوند. در پستانداران کوچکتر (مانند موشها)، نواحی عایق مغز اطلاعات خود را از مدالیتههای حسی-پیکری، مانند دیدگاه کنترلی در مورد حرکت سبیلها دریافت میکنند، زیرا سبیلها ابزار حسی و تشخیصی مهمی در این حیواناتاند.[۱۴]
از نظر کارکردی، فرض میشود که عایق مغز توجه را میان مدالیتههای گوناگون جدا میکند. توجه خود بهصورت پردازش بالا به پایین یا پایین به بالا تعریف میشود؛ هر دو نوع با ساختار و کارکرد مشاهدهشده در عایق مغز سازگار هستند و فرض بر این است که تعاملاتی با نواحی حسی سطحبالا که در رمزگذاری اشیاء و ویژگیها نقش دارند، صورت میگیرد. برای نمونه، ورودی از قشر پیشپیشانی توجه را بر پایهٔ رفتار شناختی سطحبالا و وظیفهمحور تعیین میکند. همچنین القای تحریک الکتریکی در عایق مغز باعث مهار خواندن، خیره شدن بدون واکنش و عدم پاسخدهی شده است. گزارش شده که عایق مغز دارای فرکانس پایهای خاصی برای تخلیهٔ الکتریکی است که این بسامد میتواند با توجه هدایتشده افزایش یا کاهش یابد. برای نمونه، فرافکنی به نواحی حرکتی و چشمی به حرکت نگاه کمک میکند تا توجه به سوی محرکهای تازه هدایت شود و این امر از طریق افزایش فرکانس تخلیهٔ نورونهای عایق مغز انجام میشود.[۱۴]
سالوینورین، ترکیب توهمزای اصلی موجود در گیاه سالویا دیوینوروم، میتواند موجب از دست رفتن آگاهی شود. مصرف سالوینورین A میتواند موجب همحسی (Synesthesia) شود، یعنی تفسیر مدالیتههای حسی مختلف از سوی قشرهای حسی نادرست (مثلاً شنیدن رنگها یا چشیدن صداها). این پدیده از ایدهٔ جداسازی و هدایت درونتالاموسیِ اطلاعات (توجه) پشتیبانی میکند. عایق مغز دارای گیرندهٔ افیونی کاپا است که سالوینورین A به آن متصل شده و این اثر را القا میکند.[۵][۱۴]
شواهد تجربی
تحریک با بسامد بالا (HFS) در عایق مغز گربه میتواند تغییراتی در دستگاه خودمختار ایجاد کرده و باعث بروز «نشانگان غیرفعالسازی» شود. این نشانگان با کاهش آگاهی مشخص میشود و نشاندهندهٔ ارتباط میان عایق مغز و خودآگاهی است.[۴۲] همین اثر در انسان نیز مشاهده شده است. تحریک عایق مغز چپ در انسان باعث «توقف کامل رفتار ارادی، عدم پاسخدهی، و فراموشی» شده، بیآنکه نشانههای منفی حرکتی یا صرفاً زبانپریشی ایجاد شود؛ موضوعی که نشان از نقش عایق مغز در آگاهی دارد.[۱۵] همچنین مطالعات امآرآی نشان دادهاند که افزایش شدت سیگنال درون عایق مغز با وضعیت صرع پایدار[۴۳] همراه است؛ حالتی که در آن حملههای صرعی بدون بازگشت آگاهی میان آنها روی میدهد.[۴۴][۴۵] افزایش سیگنال همچنین با حملههای موضعی-شناختی (focal dyscognitive seizures) در ارتباط است؛ این حملهها باعث اختلال در آگاهی یا هشیاری میشوند بیآنکه با تشنج همراه باشند. در این حالت، فرد از محیط اطراف خود آگاه نیست و حمله با خیرهشدن بیحرکت برای مدتی کوتاه همراه است.
در آزمایشی با استفاده از شرطیسازی فعال بههمراه HFS در عایق مغز موش، تغییرات رفتاری قابلتوجهی از جمله واکنشهای حرکتی تعدیلشده، بیحرکتی و کاهش پاسخدهی مشاهده شد.[۱۵] فراتر از این، مطالعات نشان دادهاند که عایق مغز در هنگام خواب حرکات سریع چشم فعال است؛ ساختارهایی مانند برجستگی دندانهدار نیز در این وضعیت فعالاند و نقشی وابسته در حافظه فضایی دارند، که نشان میدهد احتمالاً در این نواحی نوعی تثبیت حافظه رخ میدهد.[۶]
ضایعهها و خودآگاهی
از نظر کارکردی، عایق مغز از طریق ارتباطات خود، ورودیهای قشری گوناگون را در فرایند خودآگاهی یکپارچه میکند. بر پایهٔ ساختار و پیوندهای آن، گمان میرود که کارکرد عایق مغز با هماهنگی عملکردهای مختلف مغزی مرتبط باشد؛ همانند تمثیل «رهبر ارکستر». خودآگاهی بهطور کارکردی به دو مؤلفه تقسیم میشود: (i) بیداری، که شامل برانگیختگی و هشیاری است؛ (ii) محتوای خودآگاهی، که شامل پردازش محتوا است.
مطالعهای بر روی آسیبهای مغزی ناشی از جنگ در کهنهسربازان برای درک بهتر نقش عملکردی عایق مغز انجام شد. آسیب به عایق مغز با طول مدت بیهوشی (LOC) مرتبط بود، اما نه با تعداد دفعات آن. همچنین هرچه اندازهٔ ضایعه بزرگتر بود، طول مدت بیهوشی نیز بیشتر میشد. هیچ اثر قابلتوجهی بر پردازش شناختی مشاهده نشد.[۵]
در مطالعهای بر روی یک مورد منفرد، تحریک کپسول بسیار خارجی مغز – که در نزدیکی عایق مغز قرار دارد – باعث اختلال در خودآگاهی شد؛ بهگونهای که بلافاصله پس از قطع تحریک، آگاهی بازگشت.[۱۵] در پژوهشی دیگر دربارهٔ نشانهشناسی روانگسیختگی، مشخص شد که شدت هذیانها با کاهش حجم ماده خاکستری در عایق مغز چپ در ارتباط است؛ این موضوع نشان میدهد که میان ساختار عایق مغز و نشانههای مثبت این اختلال روانی، همبستگی وجود دارد. یافتههای دیگری این همبستگی را تأیید کردهاند، از جمله اینکه حجم ماده سفید واردشونده به عایق مغز در بیماران مبتلا به روانگسیختگی (اسکیزوفرنی) افزایش مییابد.[۴۶] همچنین، همبستگی معکوسی میان حجم ماده خاکستری و شدت توهم شنوایی در اسکیزوفرنی گزارش شده است.[۴۷] برای مشاهدهٔ فقدان کامل عملکرد عایق مغز، باید هر دو عایق مغز در دو نیمکره دچار ضایعه شوند.[۲]
با اینحال، مطالعهای در سال ۲۰۱۹ که به تحریک الکتریکی عایق مغز پرداخت، در هیچیک از پنج بیمار بررسیشده اختلالی در خودآگاهی مشاهده نکرد. این بیماران در حین آزمایش تجربیات ذهنی حسی گوناگونی گزارش کردند و حرکتهای بازتابی داشتند، اما هیچکدام از آنها دچار از دست دادن آگاهی نشدند. این یافته، نقش عایق مغز در ایجاد اختلال در آگاهی در پاسخ به تحریک الکتریکی را زیر سؤال میبرد.[۴۸]
اهمیت بالینی
روانگسیختگی
آسیب به عایق مغز میتواند علائمی مشابه با بیماریها یا اختلالات روانی رایج ایجاد کند. بهنظر میرسد رشد تأخیری این ساختار با اوتیسم مرتبط باشد. همچنین، یافتهها نشان میدهد که عایق مغز ممکن است در روانگسیختگی (اسکیزوفرنی) نقش داشته باشد، چراکه کاهش حجم ماده خاکستری در عایق مغز چپ و جزیره مغز راست با افزایش نشانههای مثبت مانند هذیان در ارتباط است.[۴۷]
صرع
نقش عایق مغز در صرع نیز مشاهده شده است؛ امآرآیها نشان دادهاند که شدت سیگنال در این ساختار در مبتلایان به صرع افزایش مییابد. در برخی موارد، بهویژه در حضور کاینیک اسید در مراحل آغازین، بهنظر میرسد که حملههای صرعی منشأیی در عایق مغز دارند.
خودآگاهی
مطالعهای موردی نشان داد که تحریک الکتریکی ناحیهٔ بین جزیره مغز و عایق مغز باعث اختلال در خودآگاهی میشود و بهمحض توقف تحریک، آگاهی بازمیگردد.[۵][۱۵] بیمارانی که در عایق مغز چپ دچار ضایعه بودند، بیش از کسانی که در نقاط دیگر مغز ضایعه داشتند، دچار بیهوشی شدهاند.[۵] برای نمونه، بیماری که الکترود به عایق مغز او متصل شده بود، خواندن را متوقف کرد، به نقطهای خیره شد و واکنشی نشان نداد؛ اما بهمحض برداشتن الکترود، دوباره خواندن را آغاز کرد و هیچگونه خاطرهای از دورهٔ بیواکنشی نداشت.[۱۴]
با این حال، در مطالعهای در سال ۲۰۱۹ که به تحریک الکتریکی عایق مغز پرداخت، هیچگونه اختلالی در خودآگاهی در هیچیک از پنج بیمار مشاهده نشد. آنها تجربیات حسی ذهنی و حرکات بازتابی را گزارش دادند، اما دچار از دست رفتن آگاهی نشدند. این یافتهها توانایی عایق مغز در ایجاد اختلال در آگاهی از طریق تحریک الکتریکی را زیر سؤال میبرد.[۴۸]
در مطالعهای در سال ۲۰۲۰، فعالسازی مصنوعی عایق مغز با استفاده از تحریک نوریِ اپتوژنتیک باعث خاموشی فعالیت مغزی در سراسر قشر مغز شد؛ پدیدهای که با نام «وضعیت پایین» (Down state) شناخته میشود و معمولاً در هنگام خواب یا بیداری آرام در موشها دیده میشود.[۴۱] نویسندگان این مطالعه میگویند: «عایق مغز هماهنگکنندهٔ فعالیت آهستهٔ سراسری در مغز است و بسیار هیجانانگیز است که در حال نزدیک شدن به کشف پیوندهای خاص بین ساختارهای مغزی و معمای بزرگ خودآگاهی هستیم.»
با این حال، استوان هارند، دانشمند علوم شناختی، معتقد نیست که عایق مغز «کلید روشن/خاموش» برای خودآگاهی باشد، بلکه تنها در بیداری نقش دارد. او استدلال میکند که اگر واقعاً چنین بود، تحریک مصنوعی عایق مغز نباید باعث از حال رفتن فرد میشد، بلکه فرد باید به رفتار عادی خود ادامه میداد و بعدها گزارش میداد که در آن مدت هیچ احساسی نداشته است.[۴۹]
سیلوسایبین
عایق مغز تراکم بالایی از گیرنده 5-HT2A دارد، به همین دلیل بهشدت تحت تأثیر روانگردانهایی مانند سیلوسایبین قرار میگیرد. بهنظر میرسد سیلوسایبین اتصال عملکردی میان عایق مغز و شبکه حالت پیشفرض (DMN) و نیز شبکهٔ کنترل وظایف پیشانی-جداری (FPTC) را تغییر میدهد. یافتهها نشان دادهاند که سیلوسایبین بهطور چشمگیری اتصال عملکردی عایق مغز راست با DMN را کاهش میدهد، و در عوض ارتباط آن را با FPTC افزایش میدهد.[۵۰]
پارکینسونیسم
تیمی از پژوهشگران به سرپرستی دانشمندان اعصاب در مرکز پزشکی بت ایزرائیل دیکونس، ضایعه در عایق مغز را بهعنوان منشأ احتمالی پارکینسونیسم در شرایط گوناگون شناسایی کردهاند. این گروه با استفاده از روشی نوین به نام «نگاشت شبکهای ضایعات» توانستند منشأ پارکینسونیسم را در ۲۹ بیماری که نشانههایشان ناشی از بیماری پارکینسون نبود بلکه بهدلیل ضایعات قابلمشاهده در تصویربرداری مغزی بود، تعیین کنند. نگاشت این ۲۹ ضایعه که در بخشهای مختلف مغز قرار داشتند، نشان داد که ارتباط با عایق مغز حساسترین و اختصاصیترین نشانگر بروز پارکینسونیسم ناشی از ضایعه است.[۵۱]
اضطراب و فشار روانی
در موشها، سرکوب فعالیت عایق مغز باعث کاهش اضطراب و فشار روانی و افزایش مقاومت در برابر استرس مزمن شده است.[۵۲]
سایر جانوران
در جانوران، از طریق ردیابی مسیرهای عصبی، مشخص شده است که عایق مغز با قشر مغز، نواحی حسی و حرکتی، و همچنین هیپوکامپ پیوندهای گستردهای دارد. در این مطالعات از مدلهای گوناگون جانوری مانند گربه، جوندگان و میمونها استفاده شده است.
_(18167720666).jpg)
گربهها
در گربهها، تحریک با بسامد بالا (HFS) در عایق مغز میتواند فعالیت حرکتی را تغییر دهد، موجب تغییرات در دستگاه عصبی خودمختار شود و نشانگان «غیرفعالسازی» ایجاد کند که با «کاهش آگاهی» توصیف میشود. ثبتهای عصبی، عمدتاً در گربهها و نخستیها، نشان دادهاند که نورونهای عایق مغز به محرکهای حسی پاسخ میدهند و همچنین در هنگام حرکات ارادی فعال میشوند.[۶] نقشهبرداری از قشر بینایی به عایق مغز تنها یک نقشه را شامل میشود که نواحی V1 و سه ناحیهٔ بینایی دیگر را در بر میگیرد. سلولهای V1 در لایهٔ ۶ قرار دارند و با سلولهایی که به لوب زانویی جانبی میروند متفاوتاند؛ این سلولها از گلوتامیک اسید بهعنوان ناقل عصبی استفاده میکنند.
عایق مغز گربه دارای سه ناحیهٔ مشخص است:
- ۱. ناحیهٔ پشتی-قدامی که با قشرهای حرکتی و حسی-پیکری مرتبط است،
- ۲. ناحیهٔ پشتی-خلفی که با قشر بینایی پیوند دارد،
- ۳. ناحیهای دیگر در زیر ناحیهٔ بینایی که با نواحی شنوایی در ارتباط است.[۲]
ورودیهای حسی بر پایهٔ مدالیتهها (دیداری، شنیداری و غیره) تفکیک شدهاند و ترجیح آشکاری برای اطلاعات حسی پیرامونی وجود دارد. در گربه، ورودیها از نواحی مختلف قشر بینایی دریافت میشوند و به همان نواحی نیز بازمیگردند. این چرخهها ماهیتی بینایینگر (رتینوتوپیک) دارند؛ به این معنا که ناحیهای از قشر مغز که به عایق مغز فرافکنی دارد، اطلاعات مربوط به همان بخش از میدان دید را پردازش میکند. عایق مغز در زمینهٔ بینایی تنها یک نقشه از نیمهٔ مقابل میدان دید دارد، و اطلاعات مربوط به حرکت در پیرامون میدان دید را دریافت میکند و گزینشپذیری خاصی ندارد.[۵۳][۵۴]
در زمینهٔ حس پیکری، عایق مغز گربه ورودیهای فراوانی از قشر حسی-پیکری نخستین (S1) دریافت میکند، اما ورودیهای آن از قشر حسی-پیکری دوم (S2) ضعیفترند. ورودیهای S1 با ورودیهایی از قشر حرکتی نخستین، بهویژه از نواحی مربوط به پنجهٔ پیشین، همپوشانی دارند.[۵۵] در مقابل، در جوندگان، عایق مغز هیچ ورودیای از S1 یا S2 دریافت نمیکند و عمدتاً تحت کنترل قشر حرکتی است.
جوندگان
در موش صحرایی، نواحی حرکتیِ سبیلها ورودی خود را از عایق مغز همسمت دریافت میکنند، اما سپس به عایق مغز نیمکرهٔ مقابل فرافکنی دارند.[۶] قشر بشکهای (نقشهٔ حسی سبیلها) و قشر بینایی نخستین نیز ورودیهایی از عایق مغز همسمت دریافت میکنند، اما تنها تعداد اندکی فرافکنی بهسمت عایق مغز دارند. این الگوها نشان میدهند که پیوندهای عایق مغز با نواحی مختلف قشری، الگوهای مشخصی دارند و نقش آن بهجای کارکردی پراکنده، بیشتر به پردازشهای تخصصی محدود میشود.[۶]
در موشها، رشتههای دارای پاروالبومین بهطور شیمیایی و الکتریکی بسیار بههم متصلاند. این رشتهها با نورونهای فرافکن عایق-قشری نیز بهشدت پیوستهاند که نشان میدهد این نورونهای بازدارنده فعالیت آنها را بهشدت تنظیم میکنند.[۶] این شبکههای محلی احتمالاً فعالیت نورونهای فرافکن عایق-قشری را هماهنگ کرده و ریتمهای مغزی و فعالیت هماهنگ نواحی مختلف قشری را تحت تأثیر قرار میدهند. ردههای دیگری از نورونهای بازدارنده نیز در عایق مغز حضور دارند که با نورونهای عایق-قشری پیوندهای محلی دارند.[۶]
آزمایشهایی که فعالیت آکسونهای عایق-قشری را در برابر محرکهای بینایی در موشها بررسی کردند، نشان دادند که عایق مغز به تغییرات محرک پاسخ میدهد.[۶] اگرچه ورودیهای عایق-قشری به نواحی بینایی درگیر بودند، قویترین پاسخها در نواحی سطحبالای قشر مغز، بهویژه در قشر کمربندی قدامی که بهطور متراکم توسط فرافکنیهای عایق مغز عصبگیری میشود، ثبت شد.[۶]
میمونها
در میمون، عایق مغز با نواحی قشر قدیم (allocortex) و قشر نو پیوندهای گستردهای دارد. این پیوندها به لوب پیشانی، نواحی قشری بینایی، قشر گیجگاهی، قشر جداری-پسسری و نواحی حسی-پیکری فرافکنی دارند.[۲] نواحی زیرقشری دریافتکنندهٔ این فرافکنیها شامل بادامک، هسته دمدار و اسبک هستند. مشخص نیست که آیا ناحیهای از قشر مغز وجود دارد که ورودی از عایق مغز نداشته باشد یا نه. در مغز میمون، نورونهای بیخار کوچک یا بزرگ نیز گزارش شدهاند که بهعنوان نورونهای مدار محلی شناخته میشوند.
عایق مغز پشتی با ساختارهای حرکتی قشر مغز پیوندهای دوسویه دارد.[۲] رابطهٔ میان حرکات جانور و رفتار نورونهای عایق مغز در ناحیهٔ پشتی-دُمی به این صورت است: ۷۰٪ نورونهای حرکتی گزینشناپذیرند و میتوانند برای هر یک از حرکات فشار، کشش یا چرخش در اندام جلویی شلیک کنند؛ بقیهٔ نورونها انتخابیتر بودند و تنها یکی از این سه حرکت را اجرا میکردند.[۲]
منابع
- ↑ شیرازی، رضا؛ اسفندیاری، ابراهیم. نورو آناتومی بالینی اسنل ۲۰۱۹ (ویراست هشتم). تهران: اندیشه رفیع. صص. ۶۷۸. شابک ۹۷۸-۹۶۴-۹۸۷۸-۸۰-۵.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Crick FC, Koch C (June 2005). "What is the function of the claustrum?". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 360 (1458): 1271–9. doi:10.1098/rstb.2005.1661. PMC 1569501. PMID 16147522.
- ↑ Smith JB, Lee AK, Jackson J (2020). "The claustrum". Current Biology. 30 (23): R1401–R1406. Bibcode:2020CBio...30R1401S. doi:10.1016/j.cub.2020.09.069. PMID 33290700. S2CID 227507231.
- 1 2 Smith JB, Alloway KD, Hof PR, Orman R, Reser DH, Watakabe A, Watson GDR (February 2019). "The relationship between the claustrum and endopiriform nucleus: A perspective towards consensus on cross-species homology". J Comp Neurol. 527 (2): 476–499. doi:10.1002/cne.24537. PMC 6421118. PMID 30225888.
- 1 2 3 4 5 6 7 Chau A, Salazar AM, Krueger F, Cristofori I, Grafman J (November 2015). "The effect of claustrum lesions on human consciousness and recovery of function". Consciousness and Cognition. 36: 256–64. doi:10.1016/j.concog.2015.06.017. PMID 26186439. S2CID 46139982.
- 1 2 3 4 5 6 7 8 9 10 11 Brown SP, Mathur BN, Olsen SR, Luppi PH, Bickford ME, Citri A (November 2017). "New Breakthroughs in Understanding the Role of Functional Interactions between the Neocortex and the Claustrum". The Journal of Neuroscience. 37 (45): 10877–10881. doi:10.1523/JNEUROSCI.1837-17.2017. PMC 5678020. PMID 29118217.
- ↑ Smith JB, Watson G, Liang Z, Liu Y, Zhang N, Alloway KD (2020). "A Role for the Claustrum in Salience Processing?". Frontiers in Neuroanatomy. 13 (64): R1401–R1406. doi:10.3389/fnana.2019.00064. PMC 6594418. PMID 31275119.
- ↑ Braak H, Braak E (1982). "Neuronal types in the claustrum of man". Anatomy and Embryology. 163 (4): 447–60. doi:10.1007/BF00305558. PMID 7091711. S2CID 7566723.
- 1 2 Nikolenko VN, Rizaeva NA, Beeraka NM, Oganesyan MV, Kudryashova VA, Dubovets AA, Borminskaya ID, Bulygin KV, Sinelnikov MY, Aliev G (July 2021). "The mystery of claustral neural circuits and recent updates on its role in neurodegenerative pathology". Behav Brain Funct. 17 (1): 8. doi:10.1186/s12993-021-00181-1. PMC 8261917. PMID 34233707.
- ↑ Mathur BN (2014). "The claustrum in review". Front Syst Neurosci. 8: 48. doi:10.3389/fnsys.2014.00048. PMC 3983483. PMID 24772070.
- 1 2 3 Smith JB, Liang Z, Watson GD, Alloway KD, Zhang N (July 2017). "Interhemispheric resting-state functional connectivity of the claustrum in the awake and anesthetized states". Brain Structure & Function. 222 (5): 2041–2058. doi:10.1007/s00429-016-1323-9. PMC 5382132. PMID 27714529.
- ↑ Stevens CF (June 2005). "Consciousness: Crick and the claustrum". Nature. 435 (7045): 1040–1. Bibcode:2005Natur.435.1040S. doi:10.1038/4351040a. PMID 15973394. S2CID 5402518.
- 1 2 3 Torgerson CM, Irimia A, Goh SY, Van Horn JD (March 2015). "The DTI connectivity of the human claustrum". Human Brain Mapping. 36 (3): 827–38. doi:10.1002/hbm.22667. PMC 4324054. PMID 25339630.
- 1 2 3 4 5 6 7 8 9 Goll Y, Atlan G, Citri A (August 2015). "Attention: the claustrum". Trends in Neurosciences. 38 (8): 486–95. doi:10.1016/j.tins.2015.05.006. PMID 26116988. S2CID 38353825.
- 1 2 3 4 5 Koubeissi MZ, Bartolomei F, Beltagy A, Picard F (August 2014). "Electrical stimulation of a small brain area reversibly disrupts consciousness". Epilepsy & Behavior. 37: 32–5. doi:10.1016/j.yebeh.2014.05.027. PMID 24967698. S2CID 8368944.
- ↑ Baizer JS, Sherwood CC, Noonan M, Hof PR (2014). "Comparative organization of the claustrum: what does structure tell us about function?". Frontiers in Systems Neuroscience. 8: 117. doi:10.3389/fnsys.2014.00117. PMC 4079070. PMID 25071474.
- ↑ Mathur BN (2014). "The claustrum in review". Frontiers in Systems Neuroscience. 8: 48. doi:10.3389/fnsys.2014.00048. PMC 3983483. PMID 24772070.
- 1 2 Edelstein LR, Denaro FJ (September 2004). "The claustrum: a historical review of its anatomy, physiology, cytochemistry and functional significance". Cellular and Molecular Biology. 50 (6): 675–702. PMID 15643691.
- ↑ Buchanan KJ, Johnson JI (May 2011). "Diversity of spatial relationships of the claustrum and insula in branches of the mammalian radiation". Annals of the New York Academy of Sciences. 1225 Suppl 1 (S1): E30-63. Bibcode:2011NYASA1225E..30B. doi:10.1111/j.1749-6632.2011.06022.x. PMID 21599698. S2CID 2245096.
- ↑ Sherk, Helen (2014). "Physiology of the Claustrum". The Claustrum. pp. 177–191. doi:10.1016/B978-0-12-404566-8.00005-2. ISBN 978-0-12-404566-8.
- ↑ Smythies, John R; Edelstein, Lawrence R; Ramachandran, V S (2014). The claustrum: structural, functional, and clinical neuroscience. Academic Press. ISBN 978-0-12-404566-8. OCLC 861211388.
- ↑ Fernandez-Miranda JC, Pathak S, Engh J, Jarbo K, Verstynen T, Yeh FC, Wang Y, Mintz A, Boada F, Schneider W, Friedlander R (August 2012). "High-definition fiber tractography of the human brain: neuroanatomical validation and neurosurgical applications". Neurosurgery. 71 (2): 430–53. doi:10.1227/NEU.0b013e3182592faa. PMID 22513841. S2CID 12867524.
- ↑ LeVay S (December 1986). "Synaptic organization of claustral and geniculate afferents to the visual cortex of the cat". The Journal of Neuroscience. 6 (12): 3564–75. doi:10.1523/JNEUROSCI.06-12-03564.1986. PMC 6568649. PMID 2432202.
- ↑ Zingg B, Hintiryan H, Gou L, Song MY, Bay M, Bienkowski MS, Foster NN, Yamashita S, Bowman I, Toga AW, Dong HW (February 2014). "Neural networks of the mouse neocortex". Cell. 156 (5): 1096–111. doi:10.1016/j.cell.2014.02.023. PMC 4169118. PMID 24581503.
- ↑ Markowitsch HJ, Irle E, Bang-Olsen R, Flindt-Egebak P (June 1984). "Claustral efferents to the cat's limbic cortex studied with retrograde and anterograde tracing techniques". Neuroscience. 12 (2): 409–25. doi:10.1016/0306-4522(84)90062-9. PMID 6462456. S2CID 21613309.
- ↑ Smith JB, Alloway KD (December 2010). "Functional specificity of claustrum connections in the rat: interhemispheric communication between specific parts of motor cortex". The Journal of Neuroscience. 30 (50): 16832–44. doi:10.1523/JNEUROSCI.4438-10.2010. PMC 3010244. PMID 21159954.
- ↑ Smith JB, Alloway KD (2014). "Interhemispheric claustral circuits coordinate sensory and motor cortical areas that regulate exploratory behaviors". Frontiers in Systems Neuroscience. 8: 93. doi:10.3389/fnsys.2014.00093. PMC 4032913. PMID 24904315.
- 1 2 3 4 خطای یادکرد: خطای یادکرد:برچسب
<ref> غیرمجاز؛ متنی برای یادکردهای با نامShelton-2022وارد نشده است. (صفحهٔ راهنما را مطالعه کنید.). - 1 2 3 4 Marriott, Brian A.; Do, Alison D.; Zahacy, Ryan; Jackson, Jesse (2021). "Topographic gradients define the projection patterns of the claustrum core and shell in mice". Journal of Comparative Neurology (به انگلیسی). 529 (7): 1607–1627. doi:10.1002/cne.25043. ISSN 0021-9967. PMC 8048916. PMID 32975316.
- ↑ Kim, Juhyun; Matney, Chanel J.; Roth, Richard H.; Brown, Solange P. (2016-01-20). [(https://www.jneurosci.org/content/36/3/773) "Synaptic Organization of the Neuronal Circuits of the Claustrum"]. Journal of Neuroscience (به انگلیسی). 36 (3): 773–784. doi:10.1523/JNEUROSCI.3643-15.2016. ISSN 0270-6474. PMC 4719014. PMID 26791208.
{{cite journal}}: Check|url=value (help) - ↑ Orman, Rena (2015-11-01). "Claustrum: a case for directional, excitatory, intrinsic connectivity in the rat". The Journal of Physiological Sciences (به انگلیسی). 65 (6): 533–544. doi:10.1007/s12576-015-0391-6. ISSN 1880-6562. PMC 10717944. PMID 26329935. S2CID 255605784.
- ↑ Tremblay, Robin; Lee, Soohyun; Rudy, Bernardo (20 July 2016). "GABAergic interneurons in the neocortex: From cellular properties to circuits". Neuron. 91 (2): 260–292. doi:10.1016/j.neuron.2016.06.033. PMC 4980915. PMID 27477017.
- ↑ Wang, Quanxin; Wang, Yun; Kuo, Hsien-Chi; Xie, Peng; Kuang, Xiuli; Hirokawa, Karla E.; Naeemi, Maitham; Yao, Shenqin; Mallory, Matt; Ouellette, Ben; Lesnar, Phil; Li, Yaoyao; Ye, Min; Chen, Chao; Xiong, Wei (2023-02-28). "Regional and cell-type-specific afferent and efferent projections of the mouse claustrum". Cell Reports. 42 (2): 112118. doi:10.1016/j.celrep.2023.112118. ISSN 2211-1247. PMC 10415534. PMID 36774552.
- 1 2 Qadir, Houman; Stewart, Brent W.; VanRyzin, Jonathan W.; Wu, Qiong; Chen, Shuo; Seminowicz, David A.; Mathur, Brian N. (2022-12-20). "The mouse claustrum synaptically connects cortical network motifs". Cell Reports. 41 (12): 111860. doi:10.1016/j.celrep.2022.111860. ISSN 2211-1247. PMC 9838879. PMID 36543121.
- ↑ Graf, Martin; Nair, Aditya; Wong, Kelly L. L.; Tang, Yanxia; Augustine, George J. (2020-07-01). [(https://www.eneuro.org/content/7/4/ENEURO.0216-20.2020) "Identification of Mouse Claustral Neuron Types Based on Their Intrinsic Electrical Properties"]. eNeuro (به انگلیسی). 7 (4). doi:10.1523/ENEURO.0216-20.2020. ISSN 2373-2822. PMC 7405070. PMID 32527746.
{{cite journal}}: Check|url=value (help) - ↑ Erwin, Sarah R; Bristow, Brianna N; Sullivan, Kaitlin E; Kendrick, Rennie M; Marriott, Brian; Wang, Lihua; Clements, Jody; Lemire, Andrew L; Jackson, Jesse; Cembrowski, Mark S (2021-08-16). Mao, Tianyi; Westbrook, Gary L; Zhang, Li I (eds.). "Spatially patterned excitatory neuron subtypes and projections of the claustrum". eLife. 10: e68967. doi:10.7554/eLife.68967. ISSN 2050-084X. PMC 8367382. PMID 34397382.
- ↑ Nair, Aditya; Teo, Yue Yang; Augustine, George J.; Graf, Martin (2023-07-11). "A functional logic for neurotransmitter corelease in the cholinergic forebrain pathway". Proceedings of the National Academy of Sciences (به انگلیسی). 120 (28): e2218830120. Bibcode:2023PNAS..12018830N. doi:10.1073/pnas.2218830120. ISSN 0027-8424. PMC 10334726. PMID 37399414.
- ↑ divisive normalization
- ↑ Grasby K, Talk A (March 2013). "The anterior claustrum and spatial reversal learning in rats". Brain Research. 1499: 43–52. doi:10.1016/j.brainres.2013.01.014. PMID 23318254. S2CID 19605350.
- ↑ Norimoto, et al. (2020). [(http://id.nii.ac.jp/1394/00001182/) "A claustrum in reptiles and its role in slow-wave sleep"]. Nature. 578 (7795): 413–418. Bibcode:2020Natur.578..413N. doi:10.1038/s41586-020-1993-6. hdl:21.11116/0000-0008-09DB-8. PMID 32051589. S2CID 256820426.
{{cite journal}}: Check|url=value (help) - 1 2 Narikiyo, et al. (2020). "The claustrum coordinates cortical slow-wave activity". Nature Neuroscience. 23 (6): 741–753. doi:10.1038/s41593-020-0625-7. PMID 32393895. S2CID 256840965.
- ↑ Gabor, Andrew J.; Peele, Talmage L. (November 1964). "Alterations of behavior following stimulation of the claustrum of the cat". Electroencephalography and Clinical Neurophysiology. 17 (5): 513–519. doi:10.1016/0013-4694(64)90181-6. PMID 14229851.
- ↑ status epilepticus
- ↑ Silva G, Jacob S, Melo C, Alves D, Costa D (June 2018). "Claustrum sign in a child with refractory status epilepticus after febrile illness: why does it happen?". Acta Neurologica Belgica. 118 (2): 303–305. doi:10.1007/s13760-017-0820-9. PMID 28741106. S2CID 32771124.
- ↑ Meletti S, Slonkova J, Mareckova I, Monti G, Specchio N, Hon P, Giovannini G, Marcian V, Chiari A, Krupa P, Pietrafusa N, Berankova D, Bar M (October 2015). "Claustrum damage and refractory status epilepticus following febrile illness". Neurology. 85 (14): 1224–32. doi:10.1212/WNL.0000000000001996. PMC 4607596. PMID 26341869.
- ↑ Shapleske J, Rossell SL, Chitnis XA, Suckling J, Simmons A, Bullmore ET, Woodruff PW, David AS (December 2002). "A computational morphometric MRI study of schizophrenia: effects of hallucinations". Cerebral Cortex. 12 (12): 1331–41. doi:10.1093/cercor/12.12.1331. PMID 12427683. S2CID 33360335.
- 1 2 Cascella NG, Gerner GJ, Fieldstone SC, Sawa A, Schretlen DJ (December 2011). "The insula-claustrum region and delusions in schizophrenia". Schizophrenia Research. 133 (1–3): 77–81. doi:10.1016/j.schres.2011.08.004. PMID 21875780. S2CID 45564142.
- 1 2 Bickel, Stephan; Parvizi, Josef (August 2019). "Electrical stimulation of the human claustrum". Epilepsy & Behavior. 97: 296–303. doi:10.1016/j.yebeh.2019.03.051. PMID 31196825. S2CID 182952015.
- ↑ [(https://generic.wordpress.soton.ac.uk/skywritings/2019/01/01/claustrum-nostrum-no-onoff-switch-for-consciousness/) "Claustrum Nostrum: No On/Off Switch for Consciousness – Skywritings"]. generic.wordpress.soton.ac.uk. Retrieved 2023-12-02.
{{cite web}}: Check|url=value (help) - ↑ Barrett, Frederick S.; Krimmel, Samuel R.; Griffiths, Roland R.; Seminowicz, David A.; Mathur, Brian N. (September 2020). "Psilocybin acutely alters the functional connectivity of the claustrum with brain networks that support perception, memory, and attention". NeuroImage. 218: 116980. doi:10.1016/j.neuroimage.2020.116980. ISSN 1095-9572. PMC 10792549. PMID 32454209.
- ↑ Joutsa J, Horn A, Hsu J, Fox MD (August 2018). "Localizing parkinsonism based on focal brain lesions". Brain. 141 (8): 2445–2456. doi:10.1093/brain/awy161. PMC 6061866. PMID 29982424.
- ↑ Niu, M. (2022). "Claustrum mediates bidirectional and reversible control of stress-induced anxiety responses". Science Advances. doi:10.1126/sciadv.abi6375. PMID 35302853.
{{cite journal}}: Text "..." ignored (help) - ↑ خطای یادکرد: خطای یادکرد:برچسب
<ref> غیرمجاز؛ متنی برای یادکردهای با نامOlson1980وارد نشده است. (صفحهٔ راهنما را مطالعه کنید.). - ↑ خطای یادکرد: خطای یادکرد:برچسب
<ref> غیرمجاز؛ متنی برای یادکردهای با نامSherk1981وارد نشده است. (صفحهٔ راهنما را مطالعه کنید.). - ↑ Smith JB, Chakrabarti S, Mowery TM, Alloway KD (2022). "Convergence of forepaw somatosensory and motor cortical projections in the striatum, claustrum, thalamus, and pontine nuclei of cats". Brain Structure and Function. 227 (1): 361–379. doi:10.1007/s00429-021-02405-6. PMID 34665323. S2CID 253984860.
- مشارکتکنندگان ویکیپدیا. «Claustrum». در دانشنامهٔ ویکیپدیای انگلیسی، بازبینیشده در ۲ مه ۲۰۲۵.