گره رانویه
| گره رانویه | |
|---|---|
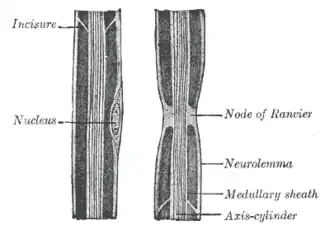 طرحی از آکسون عصب محیطی (با برچسب "axis cylinder")، که گره رانویه را به همراه سایر ویژگیها نشان میدهد. | |
 گرههای رانویه | |
| جزئیات | |
| شناسهها | |
| لاتین | incisura myelini |
| MeSH | D011901 |
| TH | H2.00.06.2.03015 |
گرههای رانویه (node of Ranvier) که به عنوان شکافهای غلاف میلین نیز شناخته میشوند، فرورفتگیهای غلاف میلین در تارهای عصبی هستند. در واقع فواصلی در رشتههای عصبی که در آنها غلاف میلین وجود ندارد، گره رانویه نام دارد. هدایت پیام عصبی در این رشته از گرهی به گره دیگر است. این گرهها شتاب پتانسیل کار از یک گره به گره بعدی را افزایش میدهد. به دلیل وجود گره رانویه در تارهای عصبی میلیندار سرعت هدایت پتانسیل کار در این تارها از سایر تارهای عصبی بیشتر است. هدایت پیام عصبی در تارهای عصبی میلین دار از یک گره رانویه به گره بعدی صورت میگیرد و به صورت جهشی است و همین امر موجب میشود که هدایت پیوسته با حداکثر سرعت ۱۲۰ متر بر ثانیه انجام شود. هدایت پیام عصبی در تارهای عصبی بدون میلین بسیار آهستهتر و با سرعت ۵ متر بر ثانیه ( حداقل سرعت) صورت میگیرند.
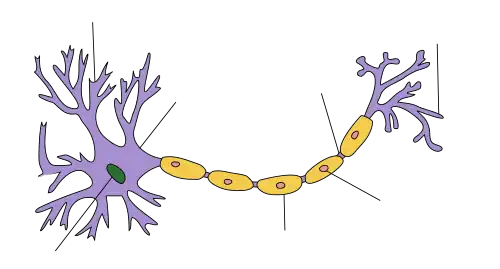
گرههای رانویه در طول یک آکسون میلیندار، در جایی که آکسولما در معرض فضای برونسلولی قرار میگیرد، رخ میدهند. گرههای رانویه دامنههای آکسونی عایقبندینشدهای هستند که سرشار از کانالهای یونی سدیمی و پتاسیمیِ متصل به مولکولهای چسبندگی سلولی میباشند و به آنها اجازه میدهند در تبادل یونهای مورد نیاز برای بازسازی پتانسیل عمل شرکت کنند.[۱] هدایت عصبی در آکسونهای میلیندار به دلیل شیوهای که به نظر میرسد پتانسیل عمل از یک گره به گره بعدی در طول آکسون «میجهد»، هدایت جهشی نامیده میشود. این امر منجر به هدایت سریعتر پتانسیل عمل میگردد. گرههای رانویه هم در دستگاه عصبی محیطی و هم در دستگاه عصبی مرکزی وجود دارند.
بررسی اجمالی
گرهها در درجه اول از کانالهای یونی دریچهدار وابسته به ولتاژ سدیمی و پتاسیمی؛ CAMها مانند نوروفاسین-۱۸۶ و NrCAM؛ و پروتئینهای آداپتور اسکلت سلولی مانند آنکیرین-G و اسپکترین βIV تشکیل شدهاند.[۲] بسیاری از آکسونهای مهرهداران توسط غلاف میلین احاطه شدهاند که امکان انتشار جهشی سریع و کارآمد پتانسیلهای عمل را فراهم میکند. تماسهای بین نورونها و سلولهای گلیال در فیبرهای میلیندار، سازماندهی فضایی و زمانی بسیار بالایی را نشان میدهند. سلولهای گلیال میلینساز — اولیگودندروسیتها در دستگاه عصبی مرکزی (CNS) و سلولهای شوان در دستگاه عصبی محیطی (PNS) — به دور آکسون پیچیده شدهاند و آکسولما را در گرههای رانویه که در فواصل منظم قرار دارند، نسبتاً بدون پوشش باقی میگذارند.
غشاهای گلیال بینگرهی برای تشکیل میلین فشرده به هم جوش خوردهاند، در حالی که حلقههای پارانودال پر از سیتوپلاسمِ سلولهای میلینساز، در هر دو طرف گرهها به صورت مارپیچی به دور آکسون پیچیده شدهاند. این سازماندهی مستلزم کنترل دقیق تکوینی و تشکیل انواع مناطق تخصصی تماس بین نواحی مختلف غشای سلول میلینساز است. هر گره رانویه توسط مناطق پارانودال احاطه شده است که در آن حلقههای گلیال که به صورت حلزونی پیچیده شدهاند، توسط اتصالی شبیه به اتصال منقطع به غشای آکسونی متصل میشوند.
بخش بین گرههای رانویه، بخش بینگرهی نامیده میشود و بیرونیترین بخش آن که با پارانودها در تماس است، به عنوان منطقه مجاور پارانودال شناخته میشود. گرهها در PNS توسط ریزپرزهایی که از جنبه بیرونی غشای سلول شوان منشأ میگیرند، یا در CNS توسط زائدههای پیرامون گرهای از آستروسیتها محصور شدهاند.
ساختار
بخشهای بینگرهی همان قطعات میلین هستند و شکافهای بین آنها گره نامیده میشوند. اندازه و فاصله بخشهای بینگرهی با قطر فیبر در یک رابطه منحنیالخط تغییر میکند که برای حداکثر سرعت هدایت بهینه شده است.[۳] اندازه گرهها از ۱ تا ۲ میکرومتر متغیر است، در حالی که بخشهای بینگرهی بسته به قطر آکسون و نوع فیبر، میتوانند تا (و گاهی حتی بیشتر از) ۱.۵ میلیمتر طول داشته باشند.
ساختار گره و مناطق پارانودال کناری با بخشهای بینگرهی زیر غلاف میلین فشرده متفاوت است، اما در CNS و PNS بسیار مشابه است. آکسون در محل گره در معرض محیط خارج سلولی قرار دارد و قطر آن منقبض شده است. کاهش اندازه آکسون منعکسکننده تراکم بستهبندی بالاتر نوروفیلامانها در این منطقه است که کمتر فسفوریله شده و با سرعت کمتری انتقال مییابند.[۴] کیسهها (وزیکولها) و سایر اندامکها نیز در گرهها افزایش مییابند که نشاندهنده وجود گلوگاه در حملونقل آکسونی در هر دو جهت و همچنین پیامرسانی موضعی آکسونی-گلیال است.
تفاوت در دستگاههای عصبی مرکزی و محیطی
اگرچه مطالعات شکست انجمادی نشان دادهاند که آکسولمای گرهای در هر دو CNS و PNS نسبت به بخش بینگرهی از ذرات درونغشایی (IMPs) غنیتر است، اما تفاوتهای ساختاری وجود دارد که منعکسکننده اجزای سلولی آنهاست.[۴] در PNS، ریزپرزهای تخصصی از یقه بیرونی سلولهای شوان بیرون زده و به آکسولمای گرهای فیبرهای بزرگ بسیار نزدیک میشوند. با این حال، در CNS، یک یا چند زائده آستروسیتی به مجاورت گرهها میآیند. پژوهشگران اعلام کردهاند که این زائدهها از آستروسیتهای چندعملکردی منشأ میگیرند. از سوی دیگر، در PNS، تیغه پایه که سلولهای شوان را احاطه کرده است، در سراسر گره پیوسته است.
ترکیبات
گرههای رانویه حاوی مبدلهای Na+
/Ca2+ و تراکم بالایی از کانالهای Na+
دریچهدار وابسته به ولتاژ هستند که پتانسیل عمل را ایجاد میکنند. یک کانال سدیمی شامل یک زیرواحد α تشکیلدهنده منفذ و دو زیرواحد کمکی β است که کانال را به اجزای خارج سلولی و داخل سلولی متصل میکنند. گرههای رانویه در دستگاههای عصبی مرکزی و محیطی عمدتاً از زیرواحدهای αNaV1.6 و β1 تشکیل شدهاند.[۵]
آنکیرین به βIV اسپکترین متصل میشود؛ ایزوفرمی از اسپکترین که در گرههای رانویه و بخشهای ابتدایی آکسون غنی است. چندین پروتئین ماتریکس خارج سلولی در گرههای رانویه متمرکز هستند، از جمله تناسین آر، Bral-1 و پروتئوگلیکان NG2. در گرههای CNS، پروتئینهای آکسونی شامل کنتاکتین نیز میشوند؛ اما برخلاف PNS، ریزپرزهای سلول شوان با زائدههای پیرامون گرهای آستروسیت جایگزین میشوند.
سازماندهی مولکولی
سطح کانالهای سدیمی در گره در مقابل بخش بینگرهی نشان میدهد که تعداد IMPها با کانالهای سدیمی مطابقت دارد. کانالهای پتاسیمی اساساً در آکسولمای گرهای غایب هستند، در حالی که در آکسولمای پارانودال و غشاهای سلول شوان در گره بسیار متمرکز هستند.[۴] عملکرد دقیق کانالهای پتاسیمی کاملاً فاش نشده است، اما دانسته شده که آنها ممکن است در دوقطبیشدن مجدد سریع پتانسیلهای عمل یا بازیابی یونهای پتاسیم در گرهها نقش حیاتی ایفا کنند.
تکوین
میلینسازی فیبرهای عصبی
تغییرات پیچیدهای که سلول شوان در طول فرآیند میلینسازی فیبرهای عصبی محیطی متحمل میشود، توسط بسیاری مطالعه شده است. پوشش اولیه آکسون بدون وقفه در تمام طول سلول شوان رخ میدهد. این فرآیند با تا خوردن سطح سلول شوان توالی مییابد تا یک غشای دو لایه از چهرههای مخالف سطح تا شده سلول شوان تشکیل شود. این غشا با ادامه تا خوردن سطح سلول شوان، بارها و بارها به صورت مارپیچی به دور خود میپیچد. در محل اتصال دو سلول شوان در طول یک آکسون، جهتهای برآمدگی لایهای انتهای میلین مخالف یکدیگر هستند.[۶] این اتصال، ناحیهای را تشکیل میدهد که به عنوان گره رانویه شناخته میشود.
مراحل اولیه
پژوهشگران ثابت کردهاند که در CNS در حال تکوین، Nav1.2 در ابتدا در تمام گرههای رانویه در حال تشکیل بیان میشود.[۷] پس از بلوغ، Nav1.2 گرهای کاهش یافته و با Nav1.6 جایگزین میشود. همچنین گزارش شده است که نوروفاسین یکی از اولین پروتئینهایی است که در گرههای رانویه تازه تشکیل شده تجمع مییابد. آنها همچنین به عنوان محل هستهگذاری برای اتصال آنکیرین G، کانالهای Nav و سایر پروتئینها عمل میکنند.
تشکیل گره
نخستین رویداد در تشکیل گره، تجمع مولکولهای چسبندگی سلول (CAMs) مانند NF186 یا NrCAM به نظر میرسد. نواحی درونسلولی این مولکولها با آنکیرین G برهمکنش دارند که به عنوان لنگری برای کانالهای سدیمی عمل میکند. در دستگاه عصبی محیطی (PNS)، این برهمکنش بهخوبی روشن شده است. پروتئین غشایی ابرخانواده Ig یعنی NrCAM، با فراخوانی آنکیرین G (یک پروتئین واسط در اتصال اسکلت سلولی اکتین-اسپکترین به کانالهای یونی موجود در گره)، به عنوان یک مولکول پیشگام در تشکیل گرهها عمل میکند.[۸][۹] همزمان، زائده پریاکسونال سلول گلیال به دور آکسون میپیچد و مناطق پارانودال را پدید میآورد. این حرکت در طول آکسون با اجازه دادن به «نیمگرههای» (heminodes) تشکیل شده در لبههای سلولهای گلیال مجاور برای ادغام و تبدیل شدن به گرههای کامل، کمک شایانی به شکلگیری کلی گرههای رانویه میکند.
اتصالات شبه-سپتات (Septate-like junctions) در پارانودها با غنیسازی NF155 در حلقههای پارانودال گلیال تشکیل میشوند.
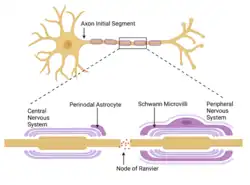
بلافاصله پس از تمایز اولیه مناطق گرهای و پارانودال، کانالهای پتاسیمی، Caspr2 و TAG1 در مناطق مجاورِ پارانودال (juxta-paranodal) تجمع مییابند. این تجمع مستقیماً با تشکیل میلین فشرده همزمان است. در مناطق گرهای بالغ، برهمکنش با پروتئینهای درونسلولی برای پایداری تمام مناطق گرهای حیاتی به نظر میرسد. در دستگاه عصبی مرکزی (CNS)، اولیگودندروسیتها فاقد ریزپرز هستند، اما به نظر میرسد قادرند از طریق فاکتورهای ترشحشده، تجمع برخی پروتئینهای آکسونی را آغاز کنند. اثرات ترکیبی چنین فاکتورهایی با حرکات بعدی ناشی از پیچش زائده اولیگودندروسیتی میتواند سازماندهی گرههای رانویه در CNS را توجیه کند.
عملکرد
پتانسیل عمل
یک پتانسیل عمل، جهشی از تخلیه یونی مثبت و منفی است که در طول غشای یک سلول حرکت میکند.[۱۰] ایجاد و هدایت پتانسیلهای عمل، ابزاری بنیادین برای ارتباط در دستگاه عصبی است. پتانسیل عمل نشاندهنده معکوس شدن سریع ولتاژ در عرض غشای پلاسمایی آکسونها است. این تغییرات سریع توسط کانالهای یونی دریچهدار وابسته به ولتاژ که در غشای پلاسمایی یافت میشوند، میانجیگری میگردند.
پتانسیل عمل از مکانی در سلول به مکان دیگر حرکت میکند، اما جریان یونی در عرض غشا تنها در گرههای رانویه رخ میدهد. در نتیجه، سیگنال پتانسیل عمل به جای انتشار نرم (آنگونه که در آکسونهای فاقد غلاف میلین رخ میدهد)، در طول آکسون از گرهای به گره دیگر میپرد. تجمع کانالهای یونی سدیمی و پتاسیمی دریچهدار وابسته به ولتاژ در گرهها، این رفتار را امکانپذیر میسازد.
هدایت جهشی
از آنجایی که یک آکسون میتواند بدون میلین یا میلیندار باشد، پتانسیل عمل دو روش برای حرکت در طول آکسون دارد. این روشها برای آکسونهای بدون میلین «هدایت پیوسته» و برای آکسونهای میلیندار «هدایت جهشی» نامیده میشوند. هدایت جهشی به صورت حرکت پتانسیل عمل در پرشهای مجزا در طول یک آکسون میلیندار تعریف میشود.
این فرآیند به این صورت است که بار بهطور غیرفعال پخش میشود تا گره رانویه بعدی را تا حد آستانه دوقطبی کند، که سپس باعث تحریک پتانسیل عمل در این منطقه میشود و دوباره بهطور غیرفعال به گره بعدی پخش میشود و این روند ادامه مییابد.
هدایت جهشی یک مزیت نسبت به هدایت در آکسونهای بدون میلین دارد و آن این است که افزایش سرعت ناشی از این حالت هدایت، تعامل سریعتر بین نورونها را تضمین میکند. از سوی دیگر، بسته به میانگین نرخ شلیک نورون، محاسبات نشان میدهد که هزینه انرژی برای حفظ پتانسیل آرامش اولیگودندروسیتها میتواند از صرفهجویی انرژی ناشی از پتانسیلهای عمل بیشتر باشد.[۱۱] بنابراین، میلیندار شدن آکسون لزوماً باعث صرفهجویی در انرژی نمیشود.
تنظیم تشکیل
تنظیم پارانود از طریق تجمع میتوکندری
میتوکندریها و سایر اندامکهای غشایی بهطور معمول در منطقه PNP آکسونهای میلیندار محیطی، بهویژه آکسونهای با قطر بزرگ، غنی هستند.[۱۲] نقش فیزیولوژیک دقیق این تجمع هنوز کاملاً درک نشده است؛ با این حال، دانسته شده که میتوکندریها معمولاً در مناطقی از سلول حضور دارند که نیاز به انرژی بالایی دارند.
در گرههای رانویه، میتوکندریها با تولید ATP که برای حفظ فعالیت پمپهای یونیِ پرمصرف ضروری است، نقش مهمی در هدایت تکانهها ایفا میکنند. در حمایت از این واقعیت، حدود پنج برابر میتوکندری بیشتری در آکسوپلاسم PNP آکسونهای محیطی بزرگ نسبت به مناطق بینگرهی مربوطه وجود دارد.[۱۲]
تنظیم گرهای
از طریق αII-اسپکترین
هدایت جهشی در آکسونهای میلیندار مستلزم سازماندهی گرههای رانویه است، جایی که کانالهای سدیمی دریچهدار وابسته به ولتاژ بسیار متراکم هستند. مطالعات نشان میدهند که αII-Spectrin (آلفا ۲ اسپکترین)، جزئی از اسکلت سلولی، در مراحل اولیه در گرهها و پارانودها غنی است و با بلوغ گرهها، بیان این مولکول ناپدید میشود.[۱۳] همچنین ثابت شده است که αII-Spectrin در اسکلت سلولی آکسونی برای تثبیت خوشههای کانال سدیمی و سازماندهی گره رانویه بالغ کاملاً حیاتی است.
تنظیم احتمالی از طریق مولکول شناسایی OMgp
پیشتر نشان داده شده بود که OMgp (گلیکوپروتئین میلین اولیگودندروسیت) در گرههای رانویه تجمع مییابد و ممکن است معماری پارانودال، طول گره و جوانهزنی آکسونی در گرهها را تنظیم کند.[۱۴] با این حال، یک مطالعه پیگیری نشان داد که آنتیبادی استفاده شده در تحقیق قبلی برای شناسایی OMgp، با یکی دیگر از اجزای غنی در گره یعنی ورسیکان V2 واکنش متقاطع (cross-react) دارد و OMgp برای یکپارچگی گرهها و پارانودها ضروری نیست.[۱۵]
اهمیت بالینی
پروتئینهای موجود در این دامنههای تحریکپذیر نورون در صورت آسیب دیدن ممکن است منجر به اختلالات شناختی و بیماریهای مختلف عصبی (نوروپاتی) شوند.
تاریخچه

غلاف میلین اعصاب بلند توسط آسیبشناس آلمانی رودولف ویرشو در سال ۱۸۵۴ کشف و نامگذاری شد.[۱۶][۱۷] آسیبشناس و کالبدشناس فرانسوی، لویی آنتوان رانویه، بعدها گرهها یا شکافهای موجود در غلاف میلین را کشف کرد که اکنون نام او را بر خود دارند. رانویه که در لیون متولد شده بود، یکی از برجستهترین بافتشناسان اواخر قرن نوزدهم بود.
رانویه این گرهها را در سال ۱۸۷۸ کشف کرد.[۱۸] او با استفاده از تکنیکهای رنگآمیزی متوجه شد که آکسونهای میلیندار تنها در فواصل منظمی رنگ میگیرند که منجر به کشف گرهها شد. گفته میشود که او در ابتدا ایده وجود گرهها در دستگاه عصبی مرکزی را رد کرد، اگرچه وجود آنها بعداً ثابت شد.[۱۹] مشاهدات او در مورد گرههای فیبر و تخریب و بازسازی فیبرهای قطعشده تأثیر زیادی بر عصبشناسی پاریس در بیمارستان سالپترییر (Salpêtrière) داشت.[۲۰]
نگارهها
 نمودار کامل سلول عصبی
نمودار کامل سلول عصبی فیبرهای عصبی مدولهشده که با نیترات نقره رنگآمیزی شدهاند
فیبرهای عصبی مدولهشده که با نیترات نقره رنگآمیزی شدهاند
منابع
- ↑ Poliak, Sebastian (2003). "The local differentiation of myelinated axons at nodes of Ranvier". Nature Reviews Neuroscience. 4 (12): 968–980. doi:10.1038/nrn1253. PMID 14682359.
- ↑ Poliak, Peles, S, E (2003). "The local differentiation of myelinated axons at nodes of Ranvier". Nat Rev Neurosci. 4 (12): 968–980. doi:10.1038/nrn1253. PMID 14682359.
- ↑ gxnSalzer J. L. (1997). "Clustering sodium channels at the node of Ranvier: close encounters of the axon-glia kind". Neuron. 18 (6): 843–846. doi:10.1016/S0896-6273(00)80323-2. PMID 9208851. S2CID 6743084.
- 1 2 3 Salzer J. L. (1997). "Clustering sodium channels at the node of Ranvier: close encounters of the axon-glia kind". Neuron. 18 (6): 843–846. doi:10.1016/S0896-6273(00)80323-2. PMID 9208851. S2CID 6743084.
- ↑ Kaplan M.R.; Cho M.H.; Ullian E.M.; Isom L.L.; Levinson S.R.; Barres B.A. (2001). "Differential control of clustering of the sodium channels Na(v)1.2 and Na(v)1.6 at developing CNS nodes of Ranvier". Neuron. 30 (1): 105–119. doi:10.1016/S0896-6273(01)00266-5. PMID 11343648. S2CID 10252129.
- ↑ Uzmman B. G.; Nogueira-Graf G. (1957). "Electron microscope studies of the formation of nodes of Ranvier in mouse sciatic nerves". Journal of Biophysical and Biochemical Cytology. 3 (4): 589–597. doi:10.1083/jcb.3.4.589. PMC 2224104. PMID 13449102.
- ↑ Boiko T, Rasband MN, Levinson SR, Caldwell JH, Mandel G, Trimmer JS, et al. (2001). "Compact myelin dictates the differential targeting of two sodium channel isoforms in the same axon". Neuron. 30 (1): 91–104. doi:10.1016/S0896-6273(01)00265-3. PMID 11343647. S2CID 7168889.
- ↑ Hill, Nishino, Nakajo, AS, A, K (2008). "Ion channel clustering at the axon initial segment and node of Ranvier evolved sequentially in early chordates". PLOS Genet. 4 (12). doi:10.1371/journal.pgen.1000317. PMC 2597720. PMID 19112491.
{{cite journal}}: Unknown parameter|article-number=ignored (help)نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Lambert, Davis, Bennett, S, JQ, V (1997). "Morphogenesis of the Node of Ranvier: Co-Clusters of Ankyrin and Ankyrin-Binding Integral Proteins Define Early Developmental Intermediates". J Neurosci. 17 (18): 7025–7036. doi:10.1523/JNEUROSCI.17-18-07025.1997. PMC 6573274. PMID 9278538.
{{cite journal}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Fry, C (2007). "Cell physiology I". Surgery (Oxford). 25 (10): 425–429. doi:10.1016/j.mpsur.2007.07.007. S2CID 57536809.
- ↑ Harris; Atwood (2012). "The Energetics of CNS White Matter". Journal of Neuroscience. 32 (1): 356–371. doi:10.1523/JNEUROSCI.3430-11.2012. PMC 3272449. PMID 22219296.
- 1 2 Einheber S, Bhat MA, Salzer JL (Aug 2006). "Disrupted Axo-Glial Junctions Result in Accumulation of Abnormal Mitochondria at Nodes of Ranvier". Neuron Glia Biology. 2 (3): 165–174. doi:10.1017/S1740925X06000275. PMC 1855224. PMID 17460780.
- ↑ Voas MG, Lyons DA, Naylor SG, Arana N, Rasband MN, Talbot WS (Mar 2007). "alphaII-spectrin is essential for assembly of the nodes of Ranvier in myelinated axons". Current Biology. 17 (6): 562–8. Bibcode:2007CBio...17..562V. doi:10.1016/j.cub.2007.01.071. PMID 17331725. S2CID 14537696.
- ↑ Huang, JK; Phillips, GR; Roth, AD; Pedraza, L; Shan, W; Belkaid, W; Mi, S; Fex-Svenningsen, A; Florens, L; Yates III, JR; Colman, DR (2005). "Glial membranes at the node of Ranvier prevent neurite outgrowth". Science. 310 (5755): 1813–17. Bibcode:2005Sci...310.1813H. doi:10.1126/science.1118313. PMID 16293723. S2CID 17410200.
- ↑ Chang, KJ; Susuki, K; Dours-Zimmermann, MT; Zimmermann, DR; Rasband, MN (2010). "Oligodendrocyte myelin glycoprotein does not influence node of Ranvier structure or assembly". J Neurosci. 30 (43): 14476–81. doi:10.1523/JNEUROSCI.1698-10.2010. PMC 2976578. PMID 20980605.
- ↑ synd/3816 در وبگاه هونِـیمدئیت؟
- ↑ Virchow R (1854). "Über das ausgebreitete Vorkommen einer dem Nervenmark analوج Substanz in den tierischen Geweben". Archiv für pathologische Anatomie und Physiologie und für klinische Medicin. 6 (4): 562–572. doi:10.1007/BF02116709. S2CID 20120269.
- ↑ Ranvier, Louis-Antoine (1871). "Contributions à l'histologie et à la physiologie des nerfs périphériques". Comptes Rendus de l'Académie des Sciences. 73.
- ↑ Tourneux, F. L. G. R., and R. Le Goff (1875). "Note sur les étranglements des tubes nerveux de la moelle épinière". Journ. De l'Anat. Et de la Phys.
{{cite journal}}: نگهداری یادکرد:نامهای متعدد:فهرست نویسندگان (link) - ↑ Barbara J.G. (2005). "Les étranglements annulaires de Louis Ranvier (1871)" (PDF). Lettre des Neurosciences. 28: 3–5.
- آرتور گایتون، جان ادوارد هال (۱۳۸۶)، «فیزیولوژی عصبی»، فیزیولوژی پزشکی گایتون، ترجمهٔ احمدرضا نیاورانی، سماط، شابک ۹۶۴-۵۷۸۹-۹۸-۲
- تونی اسمیت، سو دیویدسن (۱۳۸۴)، مغز و سیستم عصبی، ترجمهٔ عباس تیرگانی، بیژن معصوم، نشر سنبله، شابک ۹۶۴-۳۹۲-۰۷۱-۲
- پ. لاژه (۱۳۷۵)، بیولوژی و فیزیولوژی عصبی، ترجمهٔ علی حائری روحانی، انتشارات دانشگاه تهران