گیاهان C3
| بخشی از مجموعهٔ مقالات دربارهٔ |
| چرخه کربن |
|---|
 |
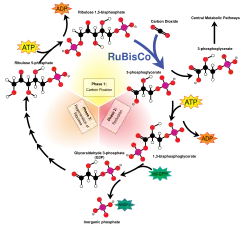
گیاهان C3 به گیاهانی گفته میشود که اولین مولکول پایدار تولیدشده از تثبیت کربن در فرایند فتوسنتزشان، دارای سه اتم کربن است. تثبیت کربن به روش C3 رایجترین مسیر از سه مسیر سوختوسازی برای تثبیت کربن در فتوسنتز است. دو مسیر دیگر C4 و CAM هستند. در این روش دیاکسید کربن و ریبولوز بیس فسفات (RuBP، یک قند ۵ کربنی) از طریق واکنش زیر به دو مولکول ۳-فسفوگلیسرات تبدیل میشود:
- CO2 + H2O + RuBP → (2) 3-phosphoglycerate
این واکنش اولین بار توسط ملوین کالوین، اندرو بنسون و جیمز باشام در سال ۱۹۵۰ کشف شد.[۱] تثبیت کربن به روش C3 در تمام گیاهان به عنوان اولین مرحله از چرخهٔ کالوین-بنسون رخ میدهد. (در گیاهان C4 و CAM، دیاکسید کربن به جای اینکه مستقیماً از هوا شود، از مالات خارج شده و وارد این واکنش میشود)

گیاهان C3 در شرایط معمولی، با شدت نور متوسط، دمای متوسط، غلظت دیاکسید کربن حدود ۲۰۰ ppm یا بالاتر، و فراوان بودن آبهای زیرزمینی، به خوبی رشد میکنند.[۲] گیاهان C3 که منشأشان به دوران مزوزوئیک و پالئوزوئیک برمیگردد، پیش از گیاهان C4 وجود داشتهاند و هنوز هم تقریباً ۹۵٪ از زیستتودهٔ گیاهی زمین، از جمله محصولات غذایی مهم مانند برنج، گندم، سویا و جو را تشکیل میدهند.
گیاهان C3 نمیتوانند در مناطق بسیار گرم با سطح CO2 اتمسفر امروزی (که در طول صدها میلیون سال از بالای ۵۰۰۰ ppm بهطور قابل توجهی کاهش یافته است) رشد کنند، زیرا روبیسکو با افزایش دما، اکسیژن بیشتری را با ریبولوز بیش فسفات ترکیب میکند. این امر منجر به تنفس نوری میشود که منجر به از دست رفتن خالص کربن و نیتروژن از گیاه میشود و بنابراین میتواند رشد را محدود کند.
گیاهان C3 تا ۹۷٪ از آبی را که از طریق ریشههایشان جذب میکنند، از طریق تعرق از دست میدهند.[۳] در مناطق خشک، گیاهان C3 روزنههای خود را میبندند تا از هدر رفتن آب جلوگیری کنند، اما این کار مانع از ورود CO2 به برگها میشود و بنابراین غلظت CO2 را در برگها کاهش میدهد. این امر نسبت CO2:O2 را کاهش میدهد و بنابراین تنفس نوری نیز افزایش پیدا میکند. گیاهان C4 و CAM سازگاریهایی دارند که به آنها اجازه میدهد در مناطق گرم و خشک زنده بمانند و بنابراین میتوانند در این مناطق از گیاهان C3 پیشی بگیرند.
امضای ایزوتوپی گیاهان C3 به دلیل تنوع تجزیهٔ ایزوتوپهای کربن در فتوسنتز اکسیژنی در انواع گیاهان، درجهٔ بالاتری از تخلیهٔ 13C را نسبت گیاهان C4 نشان میدهد. گیاهان C3 همچون گیاهان C4 پپ کربوکسیلاز ندارند که این به آنها اجازه میدهد فقط از ریبولو بیس فسفات کربوکسیلاز (روبیسکو) برای تثبیت CO2 در چرخهٔ کالوین استفاده کنند. آنزیم روبیسکو تا حد زیادی ایزوتوپهای کربن را از هم متمایز میکند و در مقایسه با 13C (ایزوتوپ سنگینتر) به ایزوتوپ 12C متصل میشود و این امر به کاهش بیشتر 13C که در C3 در مقایسه با گیاهان C4 مشاهده میشود کمک میکند، به خصوص از آنجایی که مسیر C4 علاوه بر روبیسکو از پپ کربوکسیلاز نیز استفاده میکند.[۴]
گوناگونی
همهٔ مسیرهای تثبیت کربن C3 با راندمان یکسانی عمل نمیکند.
تثبیت مجدد
بامبوها و برنجهای مرتبط با آن، راندمان C3 بهبودیافتهای دارند. این بهبود ممکن است به دلیل توانایی آن در بازجذب CO2 تولیدشده در طول تنفس نوری باشد، رفتاری که «تثبیت مجدد کربن» نامیده میشود. این گیاهان با رشد زائدههای کلروپلاست به نام «استرومول» در اطراف استروما در سلولهای مزوفیل، به تثبیت مجدد دست مییابند، بهطوری که هرگونه CO2 حاصل از تنفس نوری در میتوکندری باید از کلروپلاست پر از روبیسکو عبور کند.[۵]
تثبیت مجدد همچنین توسط طیف گستردهای از گیاهان انجام میشود. رویکرد رایج شامل رشد دستههای آوندی بزرگتر، به تنفس نوری منجر میشود.[۶]
مسیر سنتزی گلیکولات
گیاهان C3 در صورت مواجهه کمآبی مستعد تنفس نوری هستند و که این فرایند موجب جمع شدن مادهٔ سمی گلیکولات در این گیاهان میشود. دانشمندان در دههٔ ۲۰۰۰ از شبیهسازی کامپیوتری همراه با یک الگوریتم بهینهسازی استفاده کردند تا بفهمند کدام بخشهای مسیر متابولیکی ممکن است برای بهبود فتوسنتز تنظیم شوند. طبق شبیهسازی، بهبود متابولیسم گلیکولات بهطور قابل توجهی به کاهش تنفس نوری کمک میکند.[۷][۸]
ساوت و همکارانش تصمیم گرفتند به جای بهینهسازی آنزیمهای خاص در مسیر تنفس نوری برای تخریب گلیکولات، تنفس نوری را بهطور کلی نادیده بگیرند. آنها در سال ۲۰۱۹، گلیکولات دهیدروژناز کلامیدوموناس راینهاردتی و مالات سنتاز کدو تنبل بزرگ را به کلروپلاست تنباکو (یک جاندار مدل C3) منتقل کردند. این آنزیمها، به علاوهٔ آنزیمهای خود کلروپلاست، یک چرخهٔ کاتابولیک ایجاد میکنند: استیل-کوآ با گلیاکسیلات ترکیب میشود تا مالات تشکیل شود، که سپس به پیرووات و CO2 تجزیه میشود؛ پیرووات نیز به نوبهٔ خود به استیل-کوآ و CO2 تجزیه میشود. با صرف نظر کردن از تمام انتقالها بین اندامکها، تمام CO2 آزادشده صرف افزایش غلظت CO2 در کلروپلاست میشود و به تثبیت مجدد کمک میکند. نتیجهٔ نهایی، ۲۴ درصد زیستتودهٔ بیشتر است. یک جایگزین با استفاده از مسیر گلیسرات ای. کلای بهبود کمتری در حد ۱۳٪ ایجاد کرد. آنها اکنون در حال کار بر روی انتقال این بهینهسازی به سایر محصولات C3 مانند گندم هستند.[۹]
منابع
- ↑ Calvin M (1997). "Forty years of photosynthesis and related activities". Interdisciplinary Science Reviews. 22 (2): 138–148. Bibcode:1997ISRv...22..138C. doi:10.1179/isr.1997.22.2.138.
- ↑ "Respiration". Encyclopedia of Earth. Washington, D.C.: National Council for Science and the Environment. 2011.
- ↑ Raven JA, Edwards D (March 2001). "Roots: evolutionary origins and biogeochemical significance". Journal of Experimental Botany. 52 (Spec Issue): 381–401. doi:10.1093/jexbot/52.suppl_1.381. PMID 11326045.
- ↑ Alonso-Cantabrana H, von Caemmerer S (May 2016). "Carbon isotope discrimination as a diagnostic tool for C4 photosynthesis in C3-C4 intermediate species". Journal of Experimental Botany. 67 (10): 3109–21. doi:10.1093/jxb/erv555. PMC 4867892. PMID 26862154.
- ↑ Peixoto, Murilo M.; Sage, Tammy L.; Busch, Florian A.; Pacheco, Haryel D. N.; Moraes, Moemy G.; Portes, Tomás A.; Almeida, Rogério A.; Graciano-Ribeiro, Dalva; Sage, Rowan F. (27 March 2021). "Elevated efficiency of C3 photosynthesis in bamboo grasses: A possible consequence of enhanced refixation of photorespired CO2". GCB Bioenergy. 13 (6): 941–954. Bibcode:2021GCBBi..13..941P. doi:10.1111/gcbb.12819.
- ↑ Sage, Rowan F.; Khoshravesh, Roxana; Sage, Tammy L. (1 July 2014). "From proto-Kranz to C4 Kranz: building the bridge to C4 photosynthesis". Journal of Experimental Botany. 65 (13): 3341–3356. doi:10.1093/jxb/eru180. PMID 24803502.
- ↑ Zhu XG, de Sturler E, Long SP (October 2007). "Optimizing the distribution of resources between enzymes of carbon metabolism can dramatically increase photosynthetic rate: a numerical simulation using an evolutionary algorithm". Plant Physiology. 145 (2): 513–26. doi:10.1104/pp.107.103713. PMC 2048738. PMID 17720759.
- ↑ Stracquadanio G, Umeton R, Papini A, Lio P, Nicosia G (2010). "Analysis and Optimization of C3 Photosynthetic Carbon Metabolism". 2010 IEEE International Conference on BioInformatics and BioEngineering. Philadelphia, PA, USA: IEEE. pp. 44–51. doi:10.1109/BIBE.2010.17. hdl:1721.1/101094. ISBN 978-1-4244-7494-3. S2CID 5568464.
- ↑ South PF, Cavanagh AP, Liu HW, Ort DR (January 2019). "Synthetic glycolate metabolism pathways stimulate crop growth and productivity in the field". Science. 363 (6422): eaat9077. doi:10.1126/science.aat9077. PMC 7745124. PMID 30606819.