گیاهان C4
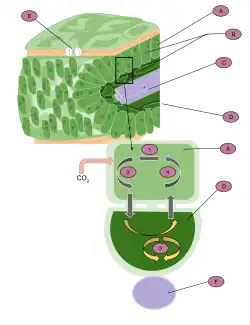
الف) سلول مزوفیل
ب: کلروپلاست
ج: بافت عروقی
د: سلول غلاف بسته
ه: استوما
و: بافت عروقی ۱. CO2 برای تولید یک مولکول چهار کربنی (مالات یا آسپارتات) تثبیت میشود
۲. این مولکول از سلول خارج شده و وارد سلولهای غلاف آوندی میشود. ۳. سپس به CO2 و پیرووات تجزیه میشود. CO2 وارد چرخه کالوین میشود و کربوهیدرات تولید میگردد.
۴. پیرووات دوباره وارد سلول مزوفیل میشود و در آنجا برای تولید مالات یا آسپارتات مورد استفادهٔ مجدد قرار میگیرد.
| بخشی از مجموعهٔ مقالات دربارهٔ |
| چرخه کربن |
|---|
 |
گیاهان C4 گروهی از گیاهان هستند که به واسطهٔ دارا بودن نوع خاصی از فتوسنتز، بهطور موثرتری در شرایط آب و هوایی گرم و خشک فعالیت میکنند. این گیاهان با استفاده از یک مکانیسم خاص، از اتلاف آب و انرژی از طریق فتوسنتز جلوگیری میکنند. تثبیت کربن به روش C4 (به انگلیسی: C4 carbon fixation) که به آن مسیر فتوسنتزی هچ اند اسلک (به انگلیسی: the Hatch–Slack pathway) نیز گفته میشود، یکی از سه فرایند فتوسنتزی شناخته شده برای تثبیت کربن در گیاهان است. این نام برگرفته از مارشال دیویدسون هچ و چارلز راجر اسلک است؛ دو دانشمندی که در دههٔ ۱۹۶۰ این مسیر فتوسنتزی را کشف کردند.[۱]
تثبیت کربن به روش C4 یک مزیت نسبت به تثبیت کربن به روش C3 دارد. آنزیم کربوکسیلهکنندهٔ اصلی در روش C3، روبیسکو نام دارد که دو واکنش متمایز را با استفاده از CO2 (واکنش کربوکسیلاسیون) یا اکسیژن (واکنش اکسیژنیشن) به عنوان سوبسترا کاتالیز میکند. اکسیژنیشن روبیسکو باعث تولید فسفوگلیکولات میشود که سمی است و برای بازیافت از طریق تنفس نوری نیاز به صرف انرژی دارد. تثبیت کربن به روش C4 با متمرکز کردن CO2 در اطراف روبیسکو، تنفس نوری را کاهش میدهد.
گیاهان C4 برای اینکه روبیسکو بتواند در محیط سلولی که دیاکسید کربن زیاد و اکسیژن بسیار کمی وجود دارد، فعالیت کند، عموماً شامل دو بخش نسبتاً جدا از هم به نام سلولهای مزوفیل و سلولهای غلاف آوندی هستند. CO2 در ابتدا در سلولهای مزوفیل در واکنشی توسط آنزیم پپ کربوکسیلاز تثبیت میشود که محصول آن فسفوانول پیرووات سه کربنی است و به دنبال آن اسید اگزالواستیک چهار کربنی تشکیل میشود. اسید اگزالواستیک بعد از این میتواند به مالات احیا شود یا به آسپارتات ترانسآمینه شود. این واسطهها به سلولهای غلاف آوندی منتشر میشوند، جایی که دکربوکسیله میشوند و محیطی غنی CO2 در اطراف روبیسکو ایجاد میکنند و در نتیجه تنفس نوری را سرکوب میکنند. پیرووات حاصل، همراه با حدود نیمی از فسفوگلیسرات تولیدشده توسط روبیسکو، به مزوفیل بازمیگردد. سپس فسفوگلیسرات به صورت شیمیایی احیا شده و دوباره به غلاف آوندی منتشر میشود تا چرخهٔ کالوین را تکمیل کند. این تبادل متابولیتها برای انجام فتوسنتز C4 ضروری است.
مراحل بیوشیمیایی اضافی برای بازسازی پپ به انرژی بیشتری به شکل ایتیپی نیاز دارند، اما غلظت بالای CO2 امکان فتوسنتز با سرعت بالا در دماهای بالاتر فراهم میکند. غلظت بالای CO2 بر کاهش حلالیت گاز با دما غلبه میکند (قانون هنری). این مکانیسم باعث میشود غلظت CO2 در منافذ روزنه نیز بالا بماند. این بدان معناست که گیاهان C4 عموماً هدایت روزنهای پایینتر، تلفات آب کمتر و عموماً راندمان مصرف آب بالاتری دارند.[۲] گیاهان C4 همچنین در استفاده از نیتروژن کارآمدتر هستند، زیرا ساخت پپ کربوکسیلاز ارزانتر از روبیسکو است.[۳] با این حال، از آنجایی که مسیر C3 برای بازسازی پپ به انرژی اضافی نیاز ندارد، در شرایطی که تنفس نوری محدود است، معمولاً در دماهای پایین و در سایه، مسیر C3 کارآمدتر است.[۴]
جستارهای وابسته
منابع
- ↑ Slack CR, Hatch MD (June 1967). "Comparative studies on the activity of carboxylases and other enzymes in relation to the new pathway of photosynthetic carbon dioxide fixation in tropical grasses". The Biochemical Journal. 103 (3): 660–5. doi:10.1042/bj1030660. PMC 1270465. PMID 4292834.
- ↑ Osborne CP, Sack L (February 2012). "Evolution of C4 plants: a new hypothesis for an interaction of CO2 and water relations mediated by plant hydraulics". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 367 (1588): 583–600. doi:10.1098/rstb.2011.0261. PMC 3248710. PMID 22232769.
- ↑ Sage RF, Pearcy RW (July 1987). "The Nitrogen Use Efficiency of C(3) and C(4) Plants: I. Leaf Nitrogen, Growth, and Biomass Partitioning in Chenopodium album (L.) and Amaranthus retroflexus (L.)". Plant Physiology. 84 (3): 954–8. doi:10.1104/pp.84.3.954. PMC 1056701. PMID 16665550.
- ↑ Bellasio C, Farquhar GD (July 2019). "3 rice: gains, losses and metabolite fluxes". The New Phytologist. 223 (1): 150–166. doi:10.1111/nph.15787. hdl:1885/159508. PMID 30859576.